¿Qué es un virus?
En biología, un virus (del latín virus, «toxina» o «veneno») es una entidad infecciosa microscópica que sólo puede multiplicarse dentro de las células de otros organismos. Los virus infectan todos los tipos de organismos, desde animales y plantas hasta bacterias y arqueas. Los virus son demasiado pequeños para poder ser observados con la ayuda de un microscopio óptico, por lo que se dice que son submicroscópicos. El primer virus conocido, el virus del mosaico del tabaco, fue descubierto por Martinus Beijerinck en 1899, y actualmente se han descrito más de 5.000, si bien algunos autores opinan que podrían existir millones de tipos diferentes. Los virus se hallan en casi todos los ecosistemas de la Tierra y son el tipo de entidad biológica más abundante. El estudio de los virus recibe el nombre de virología,6 una rama de la microbiología.
Composición.
A diferencia de los priones y viroides, los virus se componen de dos o tres partes: su material genético, que porta la información hereditaria, que puede ser ADN o de ARN; una cubierta proteica que protege a estos genes (llamada cápside) y en algunos también se puede encontrar una bicapa lipídica que los rodea cuando se encuentran fuera de la célula (denominada envoltura vírica). Los virus varían en su forma, desde simples helicoides o icosaedros hasta estructuras más complejas. El origen evolutivo de los virus aún es incierto, algunos podrían haber evolucionado a partir de plásmidos (fragmentos de ADN que se mueven entre las células), mientras que otros podrían haberse originado desde bacterias. Además, desde el punto de vista de la evolución de otras especies, los virus son un medio importante de transferencia horizontal de genes, la cual incrementa la diversidad genética.
¿Todos los virus producen enfermedades?
No todos los virus provocan enfermedades, ya que muchos virus se reproducen sin causar ningún daño al organismo infectado. Algunos virus como el VIH pueden producir infecciones permanentes o crónicas cuando el virus continúa replicándose en el cuerpo evadiendo los mecanismos de defensa del huésped. En los animales, sin embargo, es frecuente que las infecciones víricas produzcan una respuesta inmunitaria que confiere una inmunidad permanente a la infección. Los microorganismos como las bacterias también tienen defensas contra las infecciones víricas, conocidas como sistemas de restricción-modificación. Los antibióticos no tienen efecto sobre los virus, pero se han desarrollado medicamentos antivirales para tratar infecciones potencialmente mortales.
Etimología.
La palabra proviene del latín virus, que hace referencia al veneno o alguna sustancia nociva, y fue usada por primera vez en inglés en 1392. Virulento, del latín virulentus (venenoso), data del 1400. La mención de «agente que causa enfermedades infecciosa» se usó por primera vez en 1728,15 antes del descubrimiento de los virus por Dmitri Ivanovski en 1892. El adjetivo "viral" data de 1948. El término «virión» también se utiliza para referirse a una única partícula vírica infecciosa. El plural es «virus».
Origen.
Los virus son organismos parásitos que infectan células y producen viriones para difundir sus genes. La mayoría de las proteínas virales no tienen homólogos en las células modernas, en contradicción con la visión tradicional de los virus como los «ladrones de genes celulares». Esto sugiere que los genes virales básicamente tienen su origen durante la replicación de los genomas virales y/o fueron reclutados de linajes celulares ahora extintos. Algunas proteínas virales específicas están presentes en virus que infectan a los miembros de los tres dominios de la vida,39 lo que sugiere que los virus son en realidad muy antiguos. En particular, los análisis estructurales de proteínas de la cápside han revelado que al menos dos tipos de viriones se originaron de manera independiente antes que LUCA (el último antepasado universal celular). Aunque varias hipótesis han sido recientemente propuestas para explicar el origen de los virus, la aparición de viriones, como un mecanismo específico para la difusión de genes, permanece sin explicación.
Teoría de la regresión.
Es posible que los virus fueran pequeñas células que parasitaban células más grandes. A lo largo del tiempo, los genes que no necesitaban por su parasitismo desaparecieron. Las bacterias Rickettsia y Chlamydia son células vivientes que, como los virus, sólo pueden reproducirse dentro de células huéspedes. El ejemplo de estas bacterias parece apoyar esta teoría, pues es probable que su dependencia del parasitismo haya causado la pérdida de los genes que les permitían sobrevivir fuera de una célula. También se le llama «teoría de la degeneración».
Teoría del origen celular.
(También llamada «hipótesis del nomadeo» o «hipótesis del vagabundeo»): algunos virus podrían haber evolucionado a partir de fragmentos de ADN o ARN que «escaparon» de los genes de un organismo mayor. El ADN fugitivo podría haber provenido de plásmidos (fragmentos de ADN que pueden moverse entre células) o transposones. Estos son moléculas de ADN que se replican y se mueven a diferentes posiciones en el interior de los genes de la célula. Antiguamente llamados «genes saltarines», son ejemplos de elementos móviles genéticos y podrían ser el origen de algunos virus. Los transposones fueron descubiertos en 1950 por Barbara McClintock a partir de sus estudios en maíz.
Teoría de la coevolución.
Los virus podrían haber evolucionado de complejas moléculas de proteínas y ácido nucleico, al mismo tiempo que aparecieron las primeras células en la Tierra, y habrían sido dependientes de la vida celular durante muchos millones de años. Los viroides son moléculas de ARN que no son clasificadas como virus porque carecen de envoltura proteica. Sin embargo, tienen características comunes a diversos virus y a menudo se les llama agentes subvíricos. Los viroides son importantes patógenos de las plantas. No codifican proteínas, pero interactúan con la célula huésped y la utilizan para replicarse y producir sus proteínas. El virus de la hepatitis D de los humanos tiene un genoma de ARN similar al de los viroides pero tiene un envoltorio proteico derivado del virus de la hepatitis B y no puede producir uno propio. Por lo tanto, es un virus defectuoso que no puede replicarse sin la ayuda del virus de la hepatitis B. De la misma forma, el virófago 'sputnik' es dependiente del mimivirus, el cual infecta a Acanthamoeba castellanii. Estos virus que dependen de otras especies víricas reciben el nombre de satélites, y podrían representar estadios evolutivos intermedios entre los viroides y los virus.
Estructura.
Existen opiniones dispares sobre si los virus son una forma de vida o estructuras orgánicas que interactúan con los seres vivos. Por ello algunos autores se refieren a ellos como «organismos al límite de la vida». Por una parte se asemejan a los organismos que tienen genes y evolucionan por selección natural, y se reproducen creando múltiples copias de sí mismos para autoensamblarse. Sin embargo, carecen de estructura celular, lo cual es considerado la unidad básica de la vida. Además, los virus no tienen un metabolismo propio, y necesitan una célula hospedadora para crear nuevos productos. Por tanto, no se pueden reproducir en el exterior de una célula huésped (aunque bacterias como Rickettsia y Chlamydia son considerados organismos vivos a pesar de tener la misma limitación). Las formas de vida aceptadas utilizan la división celular para reproducirse, mientras que los virus aparecen de forma súbita y en gran cantidad dentro de las células, lo que es análogo al crecimiento autónomo de los cristales. El autoensamblaje de los virus dentro de las células tiene implicaciones para el estudio del origen de la vida, pues refuerza las hipótesis de que la vida podría haber comenzado en forma de moléculas orgánicas autoensamblantes.
Ácido nucleico.
El ácido nucleico en los virus contiene la información específica y el potencial para modificar operaciones en la célula infectada. Los ácidos nucleicos son macromoléculas constituidas por cadenas de nucleótidos, los cuales a su vez están constituidos por una base nitrogenada asociada a un azúcar del grupo de las pentosas y a uno o más grupos de fosfatos. Existen cuatro posibles tipos de ácido nucleico viral: ADN de cadena sencilla, ADN de cadena doble, ARN de cadena sencilla y ARN de cadena doble. Virus que contienen cualquiera de estos tipos de ácido nucleico pueden ser encontrados tanto entre los fagos como entre los virus que infectan a plantas o animales.
El ADN de algunos bacteriófagos se caracteriza por contener bases raras que substituyen alguna o algunas de las bases normalmente presentes en el ADN. El ADN de cadena doble presente en algunos virus, se caracteriza por tener segmentos de cadena sencilla en ambos extremos de la molécula. Debido a que son complementarias las secuencias de nucleótidos presentes en ambos extremos. El ADN naturalmente circular puede ser de cadena sencilla como en el fago ØXI74, o de cadena doble, como en el virus SV4O. Existe evidencia de que algunos virus ARN que producen infecciones en vegetales como el limonero y la papa contienen moléculas circulares de ARN.
En los últimos diez años se han desarrollado una variedad de técnicas y métodos que permiten determinar la secuencia de nucleótidos en cualquier tipo de ácido nucleico. La primera secuencia completa de un ARN viral fue determinada en el fago MS2 por el grupo de Walter Fiers en 1976. En 1977, Fred Sanger y colaboradores publicaron la secuencia completa del genoma del fago ØXl74, constituido por ADN de cadena sencilla. Posteriormente, muchos otros genomas virales de mayor tamaño y complejidad han sido secuenciados en parte o en su totalidad.
Estructura del virus.
Los virus presentan una amplia diversidad de formas y tamaños, llamadas «morfologías». Son unas 100 veces más pequeños que las bacterias. La mayoría de los virus estudiados tienen un diámetro de entre 10 y 300 nanómetros. Algunos Filovirus tienen un tamaño total de hasta 1.400 nm, sin embargo, sólo miden unos 80 nm de diámetro.69 La mayoría de virus no pueden ser observados con un microscopio óptico, de manera que se utilizan microscopios electrónicos de barrido y de transmisión para visualizar partículas víricas.70 Para aumentar el contraste entre los virus y el trasfondo se utilizan tinciones densas en electrones. Son soluciones de sales de metales pesados como wolframio, que dispersan electrones en las regiones cubiertas por la tinción. Cuando las partículas víricas están cubiertas por la tinción (tinción positiva), oscurecen los detalles finos. La tinción negativa evita este problema, tiñendo únicamente el trasfondo.71
Una partícula vírica completa, conocida como virión, consiste en un ácido nucleico rodeado por una capa de protección proteica llamada cápside. Las cápsides están compuestas de subunidades proteicas idénticas llamadas capsómeros.72 Los virus tienen un «envoltorio lipídico» derivado de la membrana celular del huésped. La cápside está formada por proteínas codificadas por el genoma vírico, y su forma es la base de la distinción morfológica.73 74 Las subunidades proteicas codificadas por los virus se autoensamblan para formar una cápside, generalmente necesitando la presencia del genoma viral. Sin embargo, los virus complejos codifican proteínas que contribuyen a la construcción de su cápside. Las proteínas asociadas con los ácidos nucleicos son conocidas como nucleoproteínas, y la asociación de proteínas de la cápside vírica con ácidos nucleicos víricos recibe el nombre de nucleocápside.75 76 En general, hay cuatro tipos principales de morfología vírica:
Helicoidal.
las cápsides helicoidales se componen de un único tipo de capsómero apilado alrededor de un eje central para formar una estructura helicoidal que puede tener una cavidad central o un tubo hueco. Esta formación produce viriones en forma de barra o de hilo, pueden ser cortos y muy rígidos, o largos y muy flexibles. El material genético, normalmente ARN monocatenario, pero a veces ADN monocatenario, queda unido a la hélice proteica por interacciones entre el ácido nucleico con carga negativa y la carga positiva de las proteínas. En general, la longitud de una cápside helicoidal está en relación con la longitud del ácido nucleico que contiene, y el diámetro depende del tamaño y la distribución de los capsómeros. El conocido virus del mosaico del tabaco es un ejemplo de virus helicoidal.
Icosaédrica.
la mayoría de virus que infectan los animales son icosaédricos o casi-esféricos con simetría icosaédrica. Un icosaedro regular es la mejor manera de formar una carcasa cerrada a partir de subunidades idénticas. El número mínimo requerido de capsómeros idénticos es doce, cada uno compuesto de cinco subunidades idénticas. Muchos virus, como los rotavirus, tienen más de doce capsómeros y parecen esféricos, manteniendo esta simetría. Los ápices de los capsómeros están rodeados por otros cinco capsómeros y reciben el nombre de pentones. Las caras triangulares de éstos también se componen de otros seis capsómeros y reciben el nombre de hexones.
Envoltura.
algunas especies de virus se envuelven en una forma modificada de una de las membranas celulares, o bien es la membrana externa que rodea una célula huésped infectada, o bien membranas internas como la membrana nuclear o el retículo endoplasmático, consiguiendo así una bicapa lipídica exterior conocida como envoltorio vírico. Esta membrana se rellena de proteínas codificadas por el genoma vírico y el del huésped, la membrana lipídica en sí y todos los carbohidratos presentes son codificados completamente por el huésped. El virus de la gripe y el VIH utilizan esta estrategia. La mayoría de virus envueltos dependen de la envoltura para infectar.
Complejos.
los virus tienen una cápside que no es ni puramente helicoidal, ni puramente icosaédrica, y que puede poseer estructuras adicionales como colas proteicas o una pared exterior compleja. Algunos bacteriófagos (como el Fago T4) tienen una estructura compleja que consiste en un cuerpo icosaédrico unido a una cola helicoidal (esta cola actúa como una jeringa molecular, atacando e inyectando el genoma del virus a la célula huésped), que puede tener una base hexagonal con fibras caudales proteicas que sobresalgan.
Genoma.
Se puede ver una enorme variedad de estructuras genómicas entre las especies de virus que, como grupo, contienen una diversidad genómica superior a la de los reinos de plantas, los animales o las bacterias. Hay millones de diferentes tipos de virus; y únicamente alrededor de 5.000 de ellos han sido descritos detalladamente. Un virus tiene un genoma compuesto de ADN o bien de ARN, y reciben respectivamente los nombres de «virus ADN» y «virus ARN». La gran mayoría de virus utilizan el ARN. Los virus de las plantas tienden a tener ARN monocatenario y los bacteriófagos tienden a tener ADN bicatenario.
Ciclos de virus.
La adhesión o absorción es una unión específica entre proteínas de la cápside vírica y receptores específicos de la superficie celular del huésped, pero algunos bacteriófagos también son capaces de adherirse a los flagelos, vellosidades (pili) o cápsulas presentes en la superficie de la bacteria hospedera. Para que esto suceda la bacteria debe contener el factor sexual "F" o ciertas colicinas (factores de resistencia contra agentes antimicrobianos). Los bacteriófagos filamentosos con ADN de cadena sencilla se adhieren a las puntas de estos pili mientras que los bacteriófagos esféricos de ARN se adhieren a los costados de éstos. La especificidad de unión —proteína y cápside— se determina por la variedad de huéspedes de los virus. Por ejemplo, el VIH sólo infecta linfocitos T humanos, pues su proteína de superficie, gp120, puede interactuar con la CD4 y con receptores de la superficie del linfocito T. Este mecanismo ha evolucionado para favorecer los virus que sólo pueden infectar células en que se pueden replicar. La adhesión al receptor que puede inducir cambios en la proteína de la envoltura viral que resultan en la fusión de las membranas viral y celular.
La penetración sigue a la adhesión; los virus se introducen en la célula huésped mediante endocitosis mediada por receptores o por fusión de membrana. Esto recibe a menudo el nombre de penetración vírica. La infección de las células vegetales es diferente a la de las células animales. Las plantas tienen una pared celular rígida hecha de celulosa y los virus sólo pueden entrar en las células cuando se produce un trauma en la pared celular. Los virus como el virus del mosaico del tabaco también pueden moverse directamente a las plantas, entre células, a través de poros llamados plasmodesmos. Las bacterias, como las de las plantas, tienen una fuerte pared celular que los virus tienen que romper para infectar la célula. Algunos virus han evolucionado mecanismos para inyectar su genoma a la célula bacteriana mientras la cápside viral permanece en el exterior.95
El despojo es el proceso en que la cápside vírica es degradada por enzimas virales o del huésped, liberando así el ácido nucleico del genoma vírico.
La replicación implica la síntesis de ARN mensajero (ARNm) vírico en todos los virus con rasgos de ARN positivos, la síntesis de proteínas víricas, el ensamblaje de proteínas víricas y la replicación del genoma viral. El proceso de replicación es esencial para mantener la estabilidad de la información genética contenida en el ADN. Esta replicación utiliza enzimas idénticas a las involucradas en la replicación del ADN celular y una característica común es la presencia de estructuras circulares temporales por lo menos en algunas de dicho proceso.
Tras el ensamblaje de partículas víricas, a menudo se produce una modificación postraduccional de las proteínas víricas. En virus como el VIH, esta modificación (a veces llamada «maduración»), se produce después de que el virus haya sido liberado de la célula huésped.96 El ensamblaje puede producir nuevas partículas virales. Los virus pueden autoensamblarse en un proceso similar a la cristalización, ya que las partículas virales, al igual que los cristales, constituyen estructuras que se encuentran en un estado mínimo de energía libre. Sin embargo, el genoma viral también puede especificar ciertos factores "morfogenéticos" que no contribuyen directamente a formar la estructura del virión, pero son necesarios para el proceso de ensamblaje. El fenómeno de autoensamblaje ocurre en la formación de diversas estructuras biológicas.
Los virus son liberados de la célula huésped por lisis—un proceso que mata la célula reventando su membrana. Los virus envueltos (como el VIH) son liberados de la célula huésped por gemación. Durante este proceso, el virus adquiere su envoltura, que es una parte modificada de la membrana plasmática del huésped.
Tipos de virus.
El material genético y el método por el cual los virus se replican, varían entre los diferentes tipos.
Virus ADN.
La replicación del genoma de la mayoría de virus ADN se produce en el núcleo de la célula. Si la célula tiene el receptor adecuado a la superficie, estos virus entran por fusión con la membrana celular o por endocitosis. La mayoría de virus ADN son completamente dependientes de la maquinaria de síntesis de ADN y ARN de la célula huésped, y su maquinaria de procesamiento de ARN. El genoma vírico debe atravesar la membrana nuclear de la célula para acceder a esta maquinaria.
Virus ADN bicatenario.
Este tipo de virus tiene su material genético compuesto por ADN de doble cadena y se replica usando una ADN polimerasa,98 que es dependiente del ADN y no del ARN. Este tipo de virus, por lo general, debe entrar en el núcleo de la célula huésped antes de que sea capaz de replicarse. Además, estos virus requieren de las polimerasas de la célula huésped para replicar el genoma viral y, por lo tanto, son altamente dependientes del ciclo celular.99 Para que pueda realizarse la infección y la producción de progenie del virus se requiere que la célula esté en la fase de replicación, que es cuando las polimerasas de la célula están activas. El virus puede forzar a la célula a realizar la división celular y de forma crónica esto puede conducir a la transformación de la célula y, en última instancia, producir cáncer.
Virus ADN monocatenario.
Este tipo de virus posee en su material genético ADN de cadena sencilla y se replica usando una ADN polimerasa dependiente del ADN —al igual que el Virus ADN bicatenario.102 A diferencias de los virus ADN bicatenarios, éstos poseen un ADN infectante monocatenario (de cadena simple), es decir, formado por una única cadena de nucleótidos, en lugar de la habitual doble hélice. Para que exista la replicación en este virus, es necesario que el ADN de cadena simple se convierta en ADN de cadena doble en las células infectadas.
Virus ARN.
Los virus ARN son únicos porque su información genética está codificada en ARN; esto quiere decir que usan el ácido ribonucleico (ARN) como material genético, o bien que en su proceso de replicación necesita el ARN. La replicación se suele producir en el citoplasma. Los virus ARN se pueden clasificar en unos cuatro grupos según su modo de replicación. La polaridad del ARN (si puede ser utilizado directamente o no para producir proteínas) determina en gran medida el mecanismo de replicación, y si el material genético es monocatenario o bicatenario. Los virus ARN utilizan sus propias ARN replicases para crear copias de su genoma.
Virus ARN bicatenario.
El virus ARN bicatenario es virus que posee ARN de cadena doble en su genoma. Como la mayoría de los virus ARN, se replican en el citoplasma y no dependen de las polimerasas de la células huésped como lo hacen los virus ADN, pues incluyen estas enzimas en el virión.102 La traducción suele ser monocistrónica, lo que significa que cada uno de los segmentos codifica una sola proteína, a diferencia de otros virus que exhiben una traducción más compleja. Una características partícular de éstos es su capacidad para llevar a cabo la transcripción de los segmentos de ARN bicatenarios bajo las condiciones apropiadas dentro de la cápside.
Virus ARN monocatenario positivo.
Los virus ARN monocatenarios positivos tienen ácido ribonucleico (ARN) de cadena sencilla de sentido positivo como material genético y no se replican usando ADN intermedio. Los virus ARN positivos son idénticos al ARNm viral y por lo tanto pueden ser inmediatamente traducidos por la célula huésped. Aunque el ARN purificado de un virus positivo puede causar directamente una infección, es menos infeccioso que el virus completo. La replicación tiene lugar principalmente en el citoplasma y no es tan dependiente del ciclo celular como en los virus ADN. Los virus ARN de sentido positivo tienen genomas con la misma polaridad del ARNm y pueden ser empleados directamente para la síntesis de proteínas usando la maquinaria de traducción de la célula huésped. Una de estas proteínas codificadas es la ARN replicasa, una ARN polimerasa que copia el ARN viral sin necesidad de pasar por una cadena de ADN intermedia.
Virus ARN monocatenario negativo.
Este virus tiene ácido ribonucleico (ARN) de cadena sencilla de sentido negativo como material genético y no se replica usando ADN intermedio.102 El ARN viral negativo es complementario del ARNm y por lo tanto debe convertirse en ARN positivo por una ARN polimerasa antes de la traducción. El ARN purificado de un virus negativo no es por sí mismo infeccioso puesto que necesita ser traducido en ARN positivo. Los virus ARN de sentido negativo utilizan una ARN polimerasa o transcriptasa para formar ARN de sentido positivo. Esto significa que el virus debe aportar la enzima ARN polimerasa puesto que ésta es dependiente del ARN. La molécula ARN de sentido positivo entonces actúa como un ARNm viral, que se traduce en proteínas por los ribosomas del huésped. Las proteína resultante se dedica directamente a la producción de los elementos de los nuevos viriones, tales como las proteínas de la cápsida y la ARN replicasa, que se encarga de la producción de nuevas moléculas de ARN de sentido negativo.
Virus ARN monocatenario retrotranscrito.
Un virus ARN monocatenario retrotranscrito (o virus ssRNA-RT) es un virus con ARN de cadena sencilla en su genoma que se replica en la célula huésped mediante transcripción inversa, es decir, mediante la formación de ADN a partir del molde ARN.102 Estos virus usan transcriptasa inversa codificada viralmente, es decir, una ADN polimerasa dependiente del ARN, para producir ADN a partir del genoma ARN viral. Este ADN a menudo se integra en el genoma del huésped, como en el caso de los retrovirus y seudovirus, donde es replicado y transcrito por el huésped.
Virus ADN bicatenario retrotranscrito.
Los virus de transcripción inversa se replican mediante la transcripción inversa, que es la formación de ADN a partir de una plantilla de ARN.102 Los virus de transcripción inversa que contienen un genoma de ARN utilizan un intermedio de ADN para replicarse, mientras que los que contienen un genoma de ADN utilizan un intermedio de ARN durante la replicación del genoma.
Efectos en la célula huésped.
La variedad de efectos estructurales y bioquímicos de los virus sobre las células huésped es grande. Reciben el nombre de «efectos citopáticos». La mayoría de infecciones víricas acaban provocando la muerte de la célula huésped, entre cuyas causas están la lisis de la célula, las alteraciones de la membrana superficial de la célula y la apoptosis. A menudo, la muerte de la célula es causada por el paro de sus actividades normales debido a la supresión por proteínas específicas del virus, que no son todas componentes de la partícula vírica.
Algunos virus no causan cambios aparentes en la célula infectada. Las células en que los virus es latente e inactivo presentan pocos signos de infección y a menudo funcionan normalmente. Esto causa infecciones persistentes y el virus a menudo permanece durmiente durante muchos meses o años. Este suele ser el caso del herpes simple. Algunos virus, como el virus de Epstein-Barr, a menudo hacen proliferar las células sin causar malignidad, pero otros, como los papilomavirus, son una causa demostrada de cáncer.
Clasificación.
Las clasificaciones intentan describir la diversidad de virus dándoles nombre y agrupándolos según sus semejanzas. En 1962, André Lwoff, Robert Horne y Paul Tournier fueron los primeros en desarrollar una forma de clasificación de los virus, basada en el sistema jerárquico linneano. Este sistema basa la clasificación en filos, clases, órdenes, familias, géneros y especies. Los virus fueron agrupados según sus propiedades compartidas (no las de sus huéspedes) y el tipo de ácido nucleico del que se compone su genoma. Posteriormente se formó Comité Internacional de Taxonomía de Virus.
Prevención.
Dado que los virus utilizan la maquinaria de una célula huésped para reproducirse y residen en el interior, son difíciles de eliminar sin matar la célula huésped. Los enfoques médicos más eficientes para enfrentarse a las enfermedades víricas conocidos hasta ahora son las vacunas, que ofrecen resistencia a la infección, y los antivirales.
Infección en otras especies.
Los virus infectan todo tipo de vida celular y, aunque los virus existen en todo el mundo, cada especie celular tiene un grupo de virus específico, que a menudo sólo infectan esta especie. Los virus son importantes patógenos del ganado. Enfermedades como la fiebre aftosa y la lengua azul son causadas por virus. Los animales de compañía (como perros, gatos y caballos), si no se les vacuna, son susceptibles a infecciones víricas graves. El parvovirus canino es causado por un pequeño virus ADN y las infecciones a menudo son fatales en los cachorros. Como todos los invertebrados, la abeja de la miel es susceptible a muchas infecciones víricas. Afortunadamente, la mayoría de virus coexisten de manera inofensiva con su huésped y no causan signos o síntomas de enfermedad.
Plantas.
Hay muchos tipos de virus de las plantas, pero a menudo sólo causan una pérdida de producción, y no es económicamente viable intentar controlarlos. Los virus de las plantas a menudo son transmitidos de una planta a otra por organismos conocidos como vectores. Normalmente son insectos, pero también se ha demostrado que algunos hongos, nemátodos y organismos unicelulares son vectores. Cuando se considera económico el control de infecciones por fitovirus (en los frutos perennes, por ejemplo), los esfuerzos se concentran en matar a los vectores y eliminar huéspedes alternativos como malas hierbas. Los fitovirus son inofensivos para los humanos y demás animales, pues sólo se pueden reproducir en células vegetales vivas.
Las plantas tienen mecanismos de defensa elaborados y eficientes contra los virus. Uno de los más eficientes es la presencia de los llamados genes de resistencia (R). Cada gen R confiere resistencia a un virus determinado desencadenando zonas localizadas de muerte celular alrededor de la célula infectada, que se pueden ver a simple vista en forma de manchas grandes. Esto detiene la expansión de la infección. La interferencia del ARN también es una defensa efectiva en las plantas. Cuando están infectadas, las plantas a menudo producen desinfectantes naturales que matan los virus, como el ácido salicílico, el óxido nítrico y moléculas reactivas de oxígeno.
Bacterias.
Los bacteriófagos son un grupo extremadamente común y diverso de virus. Por ejemplo, los bacteriófagos son la forma más común de entidad biológica en los medios acuáticos; en los océanos hay hasta diez veces más de estos virus que de bacterias, alcanzando niveles de 250 millones de bacteriófagos por milímetro cúbico de agua marina. Estos virus infectan bacterias específicas uniéndose a moléculas receptoras de superficie y entrando en la célula. En un periodo corto de tiempo (en algunos casos en unos minutos), las polimerasas bacterianas empiezan a traducir ARN vírico en proteína. Estas proteínas se convierten en nuevos viriones dentro de la célula, proteínas colaboradoras que ayudan a parecerse nuevos viriones, o proteínas implicadas en la lisis celular. Los enzimas víricos colaboran en la destrucción de la membrana celular y, en el caso del fago T4, un poco más de veinte minutos después de la inyección ya se pueden liberar más de 300 fagos.
El mecanismo principal por el que las bacterias se defienden de los bacteriófagos es produciendo enzimas que destruyen el ADN ajeno. Estas enzimas, llamadas endonucleases de restricción, fragmentan el ADN vírico que los bacteriófagos introducen en las células bacterianas. Las bacterias también tienen un sistema que utiliza secuencias CRISPR para retener fragmentos del genoma de virus que se han encontrado en el pasado, lo que les permite impedir la replicación del virus mediante una forma de interferencia del ARN. Este sistema genético proporciona a las bacterias una inmunidad adquirida a las infecciones.
Archea.
Algunos virus se replican dentro de los arqueas, se trata de virus ADN que parecen no tener relación con ninguna otra forma de virus y que tienen una variedad de formas inusuales, como botellas, barras con un gancho o incluso lágrimas. Estos virus han sido estudiados en mayor detalle en los termófilos, en particular los órdenes Sulfolobales y Thermoproteales. La defensa contra estos virus pueden incluir la interferencia del ARN de secuencias de secuencias repetidas de ADN del genoma arqueobacterial que están relacionadas con los genes de los virus.
Ciencias de la vida y medicina.
Los virus son importantes para el estudio de la biología molecular y celular, pues son sistemas sencillos que se pueden utilizar para manipular e investigar el funcionamiento de las células. El estudio y el uso de los virus ha ofrecido información valiosa sobre aspectos de la biología celular. Por ejemplo, los virus han resultado útiles en el estudio de la genética y han contribuido a comprender los mecanismos básicos de la genética molecular, como la replicación del ADN, la transcripción, la maduración del ARN, la traducción, el transporte de proteínas y la inmunología.
Los genetistas a menudo utilizan virus como vectores para introducir genes en células que están estudiando. Esto es útil para hacer que la célula produzca una sustancia ajena, o para estudiar el efecto de la introducción de un nuevo gen en el genoma. A este proceso se le denomina transducción. De manera similar, la viroterapia utiliza virus como vectores para tratar diversas enfermedades, pues pueden dirigirse específicamente a células y al ADN. Tiene un uso prometedor en el tratamiento del cáncer y en la terapia génica. Científicos del este de Europa han utilizado la terapia fágica como alternativa a los antibióticos desde hace un tiempo, y el interés por este enfoque está creciendo debido al alto nivel de resistencia a los antibióticos observado actualmente en algunas bacterias patógenas.
Nanotecnología.
Debido a su tamaño, forma y estructuras químicas bien definidas, los virus han sido utilizados como moldes para organizar materiales a nanoescala. Ejemplos recientes incluyen el trabajo hecho en el Naval Research Laboratory de Washington D. C., utilizando partículas del Cowpea mosaic virus (CPMV) para amplificar señales en sensores basados en chips de ADN. En este uso, las partículas víricas separan las tinciones fluorescentes utilizadas con el fin de evitar la formación de dímeros no fluorescentes que actúen como extintores. Otro ejemplo es el uso del CPMV como nanoplaca de pruebas para moléculas electrónicas.
Armas.
La capacidad de los virus de causar epidemias devastadoras en las sociedades humanas ha levantado la preocupación que se puedan convertir en armas biológicas. La preocupación aumentó después de que se consiguiera recrear el infame virus de la gripe española en un laboratorio. El virus de la viruela devastó numerosas sociedades a lo largo de la historia antes de ser erradicado. Actualmente sólo existe en varios laboratorios seguros en diversos lugares del mundo, pero los temores que pueda ser utilizado como arma no están totalmente infundados; la vacuna de la viruela no es segura —durante los años anteriores a la erradicación de la viruela, cayó más gente gravemente enferma como resultado de la vacunación que por la propia viruela — y la vacunación para la viruela ya no se practica. Por este motivo, gran parte de la población humana actual casi no tiene resistencia a la viruela.
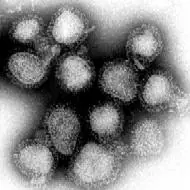
Virus de la Influenza.
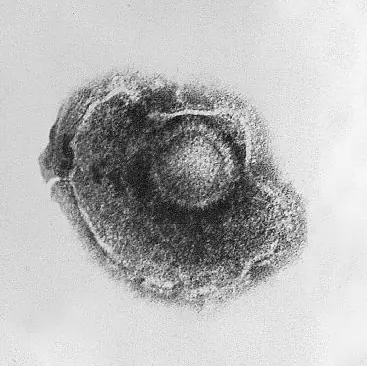
Virus del Herpes.
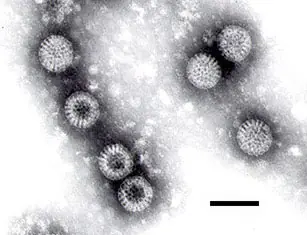
Rotavirus (tamaño de la barra 100 nm).
Fuente.
Información que tal vez te pueda interesar.