MONOGRAFIA SOBRE FOTOSINTESIS
INTRODUCCION
El diccionario define la fotosíntesis como la síntesis de un cuerpo químico en presencia de la luz solar, por la acción de la clorofila.
Ampliamente la fotosíntesis es uno de los procesos metabólicos que usan las células para obtener energía. Es en pocas palabras el único mecanismo del que dispone el mundo viviente para la producción de energía utilizable. Un proceso, mediante el cual los seres vivos que poseen clorofila, (como las plantas verdes, algas y algunas bacterias) y otros pigmentos, (como los carotenos y las xantofilas) captan energía luminosa procedente del sol (en forma de cuantos y fotones) y la transforman en energía química (ATP) y en compuestos reductores (NADPH), y con ellos transforman el agua y el CO2 en compuestos orgánicos reducidos (glucosa y otros), liberando oxígeno.
En la fotosíntesis podemos encontrar 2 etapas:
•FASE LUMINOSA se realiza en la tilacoide y se producen transferencias de electrones
•FASE OSCURA tiene lugar en el estroma y en ella se realiza la fijación de carbono
Todos los organismos que son capaces de realizar la fotosíntesis producen sus propio alimento, es decir, les permite obtener la materia y la energía que necesitan para desarrollar sus funciones vitales.
Una ecuación generalizada de la fotosíntesis se podría presentar así:
CO2 + H2O + energía -> (CH2O)n + O2
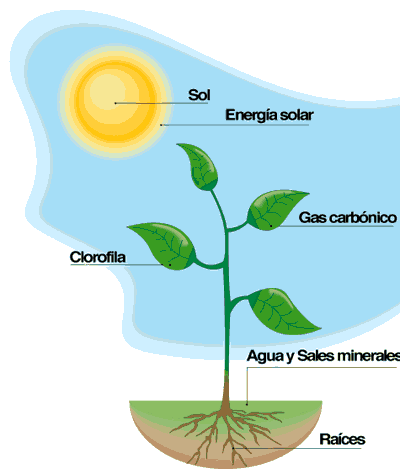
Importancia de la Fotosintesis
La fotosíntesis es seguramente el proceso bioquímico más importante de la Biosfera por varios motivos:
•La vida en la Tierra está casi completamente impulsada por el sol, y prácticamente todos los organismos dependen en última instancia del alimento producido por medio de la fotosíntesis, la cual utiliza la energía solar.
•Es responsable de la producción de toda la materia orgánica que conocemos.
•Proporciona el suministro de alimentos a prácticamente todos los seres vivos, plantas, animales, protistas, hongos y bacterias
•Produce la transformación de la energía luminosa en energía química, necesaria y utilizada por los seres vivos
•En la fotosíntesis se libera oxígeno, que se usa en la respiración aerobia como oxidante.
•La fotosíntesis fue causante del cambio producido en la atmósfera primitiva, que era anaerobia y reductora.
•De la fotosíntesis depende también la energía almacenada en combustibles fósiles como carbón, petróleo y gas natural.
•Gracias a estos procesos naturales efectuados por la planta se mantiene el equilibrio de las cantidades de dióxido de carbono y oxígeno que flotan en la atmósfera.
Ya que todos los procesos dependen unos de los otros, se puede concluir que la diversidad de la vida existente en la Tierra depende principalmente de la fotosíntesis.
Hoy día, en un mundo que tiene tanta contaminación, el aporte más importante de las plantas (en este caso de la Fotosíntesis) es la purificación del aire al termino del proceso, ya que en él, la planta despide oxígeno hacia la atmósfera que antes se encontraba en la misma como dióxido de carbono, así limpiando un poco la contaminación ambiental, provocada por humo, smog, tóxicos, etc.
FOTOSINTESIS
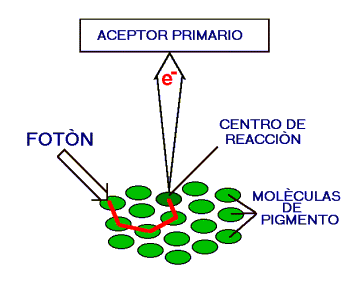
La fotosíntesis tiene lugar dentro de las celulas, en orgánulos llamados cloroplastos que contienen las clorofilas y otros compuestos, en especial enzimas, necesarios para realizar las distintas reacciones. Estos compuestos están organizados en unidades de cloroplastos llamadas tilacoides; en el interior de éstos, los pigmentos se disponen en subunidades llamadas fotosistemas. Cuando los pigmentos absorben luz, sus electrones ocupan niveles energéticos más altos, y transfieren la energía a un tipo especial de clorofila llamado centro de reacción.
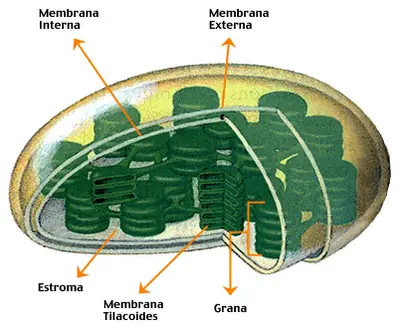
Estructura del Cloroplasto y de las membranas fotosinteticas
La unidad estructural de la fotosíntesis es el cloroplasto. Los organismos fotosintéticos procariotes y eucariotes poseen sacos aplanados o vesículas llamadas tilacoides, que contienen los pigmentos fotosintéticos; pero solamente los cloroplastos de los eucariotes están rodeados por una doble membrana. Los tilacoides se disponen como una pila de panquecas, que recibe el nombre de grana. El interior del cloroplasto entre las granas es el estroma proteico, donde se encuentran las enzimas que catalizan la fijación del CO2 . Las mitocondrias constituyen un sistema con dos membranas como los cloroplastos, pero los cloroplastos tienen tres compartimentos: el estroma, el espacio tilacoidal y el espacio entre las membranas. El cloroplasto en su interior tiene un ADN circular y ribosomas.
Clorofila y pigmentos accesorios
Un pigmento es cualquier sustancia que absorbe luz. El color de un pigmento es el resultado de la longitud de onda reflejada (no absorbida ). La clorofila, el pigmento verde de todas las células fotosintéticas, absorbe todas las longitudes de onda de la luz visible excepto el verde, el cual es reflejado y percibido por nuestros ojos.
La clorofila es una molécula compleja, formada por cuatro anillos pirrólicos, un átomo de magnesio y una cadena de fitol larga (C20H39OH).
En las plantas y otros organismos fotosintéticos existen diferentes tipos de clorofilas. La clorofila a se encuentra en todos los organismos fotosintéticos (plantas, ciertos protistas, proclorobacterias y cianobacterias). Los pigmentos accesorios absorben energía que la clorofila es incapaz de absorber. Los pigmentos accesorios incluyen clorofila b (en algas y protistas las clorofilas c,d y e), xantofila(amarilla) y caroteno, anaranjado (como el beta caroteno, un precursor de la vitamina A). La clorofila a absorbe las longitudes de ondas violeta, azul, anaranjado- rojizo, rojo y pocas radiaciones de las longitudes de onda intermedias ( verde-amarillo-anaranjado ).
Los pigmentos accesorios actúan como antena, conduciendo la energía que absorben hacia el centro de reacción. Una molécula de clorofila en el centro de reacción puede transferir su excitación como energía útil en reacciones de biosíntesis.
Cuando la clorofila absorbe energía luminosa pueden ocurrir tres cosas:
1.Que la energía sea atrapada y convertida en energía química como en la fotosíntesis
2.Que se disipe como calor
3.Que sea emitida inmediatamente como una longitud de onda mayor con perdida de energía como fluorescencia.
Fases de la fotosintesis
La fotosíntesis es un proceso que ocurre en dos fases. La primera fase es un proceso que depende de la luz (reacciones luminosas), requiere la energía directa de la luz que genera los transportadores que son utilizados en la segunda fase. La fase independiente de la luz (reacciones de oscuridad), se realiza cuando los productos de las reacciones de luz son utilizados para formar enlaces covalentes carbono-carbono (C-C), de los carbohidratos. Las reacciones oscuras pueden realizarse en la oscuridad, con la condición de que la fuente de energía (ATP) y el poder reductor (NADPH) formados en la luz se encuentren presentes. Las reacciones de oscuridad se efectúan en el estroma; mientras que las de luz ocurren en los tilacoides.
Tradicionalmente, a la primera fase se le denominaba fase luminosa y a la segunda fase oscura de la fotosíntesis. Sin embargo, la denominación como "fase oscura" de la segunda etapa es incorrecta, porque actualmente se conoce que los procesos que la llevan a cabo solo ocurren en condiciones de iluminación. Es más preciso referirse a ella como fase de fijación del dióxido de carbono (Ciclo de Calvin) y a la primera como "fase fotoquímica" o reacción de Hill.
En la fase luminosa o fotoquímica, la energía de la luz captada por los pigmentos fotosintéticos unidos a proteínas y organizados en los denominados “fotosistemas" (ver más adelante), produce la descomposición del agua, liberando electrones que circulan a través de moléculas transportadoras para llegar hasta un aceptor final (NADP+) capaz de mediar en la transformación del CO2 atmosférico (o disuelto en el agua en sistemas acuáticos) en materia orgánica. Este proceso luminoso está también acoplado a la formación de moléculas que funcionan como intercambiadores de energía en las células (ATP). La formación de ATP es necesaria también para la fijación del CO2.
Fotosistemas:
Los Fotosistemas que intervienen en la fotosintesis son dos y los podriamos definir como el conjunto de proteinas transmembranales que engloban a los pigmentos fotosinteticos cuya funcion es captar la energia luminosa para utilizarla durante el transporte de electrones que servira para sintetizar moleculas energeticas. Se encuentran localizados en los tilacoides.
Fotosistema I: Capta la energia luminosa de longitud de onda inferior o igual a 700 nm. En él, encontramos diferentes pigmentos:
Pigmentos antena: Solo sirven para captar la luz y no inician ningun tipo de reaccion. En las plantas superiores, predomina la clorofila a sobre la b que se encuentra en una menor proporcion.
Centro de reaccion: Volvemos a encontrar clorofila a, pero en este caso es la molecula diana (clorofila al). Absorbe en la longitud de onda comprendida entorno a los 700 nm y es por ello que tambien recibe el nombre de clorofila P700. Encontramos un aceptor primario de electrones denominado A0 y un donador que es la plastocianina (PC).
Fotosistema II: Capta la energia luminosa de longitud de onda igual o menor a 680 nm.
Pigmentos antena: Funcionan igual que en el caso del fotosistema I, las plantas superiores en este caso presentan clorofila a, clorofila b en mayor cantidad que en el uno y tambien xantofilas.
Centro de reaccion: Aqui la molecula diana es la clorofila aII. Como absorbe a 680 nm, a dicha clorofila se la denomina tambien clorofila P680. El aceptor primario de electrones en este caso es la feofitina (Pheo) y el donador primario recibe el nombre de donador Z.
Cada uno de estos fotosistemas se encuentra asociado a polipeptidos en la membrana tilacoidal y absorben energía luminosa independientemente. En el fotosistema II, se produce la fotólisis del agua y la liberación de oxígeno; sin embargo ambos fotosistemas operan en serie, transportando electrones, a través de una cadena transportadora de electrones. En el fotosistema I se transfieren dos electrones a la molécula de NADP+ y se forma NADPH, en el lado de la membrana tilacoidal que mira hacia el estroma.
1. Etapa Clara:
Fragmentación del agua
Se empieza con la absorción de un fotón captado por el sistema de captación de luz del fotosistema II.
El centro MN, una agrupación de cuatro átomos de Mn unida a proteína, tiene capacidad de cuatro estados de oxidación, denominados S0 a S4.
El Mn cede cuatro electrones de manera secuencial cuando cuatro fotones son captados por el fotosistema II, retirando los electrones de los oxígenos del agua unida y liberando O2.
El centro permite que un electrón a la vez pase con dirección a P680.
El fotón es conducido a una clorofila, llamada centro de reacción, denominada P680.
La excitación del P680 hace pasar a la molécula del estado basal a un estado excitado de -0.8 voltios.
El P680 excitado pasa a ser un agente reductor, capaz de transferir rápidamente un electrón desde el P680 a un aceptor electrónico primario de menor energía, la feofitina a feofitina aceptora (Ph).
Las feofitinas son moléculas idénticas a las clorofilas, excepto que dos protones sustituyen al ion Mg central.
El electrón se transfiere a continuación a una serie de moléculas de plastoquinona (QA y QB) asociadas con proteínas del PSII.
Dos electrones y dos protones son captados por la plastoquinona QB; los protones son procedentes del estroma.
La plastoquinona reducida, QH2 (plastoquinol) se libera a la porción lipídica de la membrana tilacoide.
•El plastoquinol interacciona con un complejo de citocromos y proteínas Fe-S, el citocromo bf (cit bf), ligado a la membrana. Cataliza la transferencia de los electrones a una cuproproteína, la plastocianina (PC).
•Cuando el plastoquinol se oxida para producir de nuevo plastoquinona, los dos protones se liberan a la luz del tilacoide.
•La plastocianina, es una proteína móvil de la luz del tilacoide, pasa los electrones a los centros de reacción P700.
•En este proceso, el cobre de la plastocianina se reduce primero a Cu(I) y luego se reoxida a Cu(II).
•Hasta ahora se ha dejado el centro de reacción P680 con un déficit de electrones, o mejor dicho, oxidado a un oxidante fuerte, el P680+.
•El fotosistema II extrae los electrones del agua, y los transfiere al fotosistema I liberado O2.
Producción de NADPH
•El fotosistema I contiene una clorofila centro de reacción, P700, y puede absorber la luz de hasta P700.
•La excitación por un fotón absorbido por las clorofilas antena asciende los electrones del P700 desde un estado basal a un estado excitado aproximadamente de -1.3 voltios.
•Cada electrón excitado pasa a través de una cadena de transporte electrónico.
•Primero es captado por un aceptor clorofílico (A0), luego se transfiere a una molécula de filoquinona (A1, llamada también vitamina K1).
•Por ultimo es transportado por una serie de tres Fe-S (Fx, FB y FA).
•Estas proteínas contienen grupos Fe-S (inciso d).
•Por ultimo el electrón se transfiere a otra proteína Fe-S, la ferredoxina soluble (Fd), que se encuentra en el estroma.
•La enzima ferredixina: NADP+ oxidorreductasa cataliza la transferencia de electrones al NADP+, una vez que la ferredoxina ha sido reducida por el fotosistema I:
•Es la ferredoxina, y no el NADP+, la que puede considerarse el receptor directo de los electrones de la ruta.
•Gran parte de la ferredoxina reducida se utiliza para reducir el NADP+, una gran parte se emplea para otras reacciones reductoras.
•Se puede considerar a la ferredixina reducida como una fuente de electrones de bajo potencial para mucha procesos reductores.
•El NADPH producido por la oxidación de la ferredoxina se libera al estroma, en donde se utilizara en las reacciones oscuras.
•Los electrones que se han conducido a través del fotosistema I tenían su origen en la transferencia de electrones desde los centros de reacción P700.
•Los centros de reacción oxidados (P700+) producidos de esta forma deben recibir un nuevo aporte de electrones para que continué la fotosíntesis.
•En la fotosíntesis de dos sistemas, estos electrones los proporciona el fotosistema II a través de la plastocianina.
Durante la fase luminosa aciclica tiene lugar la sintesis tanto de Atp como de NADPH + H+. Ahora bien, la cantidad de ATP sintetizada es escasa e insuficiente para la fase biosintetica dado que se necesitan 3 ATP por cada 2 NADPH y hemos acabado la fase con 2 ATP y 2 NADPH.
La fase luminosa ciclica es importante para sintetizar el ATP suficiente para que tenga lugar la fase biosintetica.
Reaccion Global:
H2O + NADP+ + Pi + ADP+ cloroplastos + luz ===>½ O2 + NADPH + H+ + ATP + H2 O
2. Etapa oscura:
Las reacciones independientes de la luz (mal llamadas reacciones oscuras) son reacciones químicas que convierten el dióxido de carbono y otros compuestos en glucosa Estas reacciones no requieren la luz para producirse (de ahí el nombre de reacciones oscuras). Estas reacciones toman los productos de la reaccion luminosa (principalmente el ATP y el NADPH) y realizan más procesos químicos sobre ellos. Las reacciones oscuras son dos: la fijacion del Carbono y el Ciclo de Calvin.
El anhídrido carbónico de la atmósfera (o del agua en los organismos acuáticos) es capturado y modificado por la adición de hidrógeno para formar carbohidratos. ([CH2O]n ). La transformación del anhídrido carbónico en un compuesto orgánico se conoce como fijacion del Carbono. La energía para ello proviene de la primera fase de la fotosíntesis.
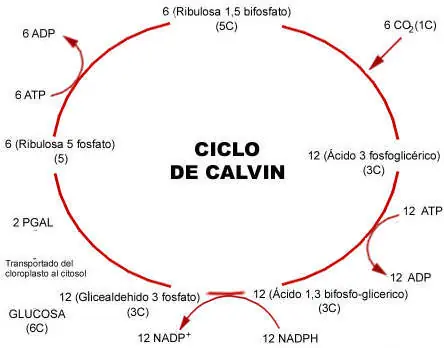
Ciclo de Calvin: Ciclo de tres fases
1.Fase de fijación del CO2: la RubisCO cataliza la reacción entre la ribulosa bifosfato (una pentosa, es decir un azúcar de 5C, RuBP) con el CO2, para crear 1 molécula de 6 carbonos, la cual al ser inestable termina por separarse en 2 moléculas que contienen 3 átomos de carbono, PGA (Fosfoglicerato). La importancia de la RuBisCo queda indicada por el hecho de ser la proteína más abundante en la naturaleza.
2.Fase de reducción: primero ocurre un proceso de activación en el cual una molécula de ATP, proveniente de la fase fotoquimica, es usada para la fosforilización del PGA, transformándolo en difosfoglicerato. Esa transferencia de un enlace fosfato permite que una molécula de NADPH+H+ reduzca el PGA, mediante la acción de la enzima gliceraldehido - 3-fosfato-deshidrogenasa, para formar gliceraldehido-3-fosfato (PGAL). Esta última molécula es una triosa-fosfato, es decir un azucar de tipo aldosa con 3C, que es una molécula estable y con mayor energía libre (capaz de realizar mayor cantidad de trabajo) que las anteriores.
Parte de PGAL se transforma en su isomero dihidroxiacetona fosfato (cetosa de 3C). Estas dos triosas-fosfato serán la base a partir de la cual se formen el resto de azúcares (como la fructosa y glucosa), oligosacáridos (como la sacarosa o azúcar de caña) y polisacáridos (como la celulosa o el almidón). También, a partir de estos azúcares se formarán directa o indirectamente las cadenas de carbono que componen el resto de moléculas que constituyen los seres vivos (lípidos, proteínas, ácidos nucleícos y otros).
3.Fase de regeneración: el ciclo continua a lo largo de una serie de reacciones hasta formar ribulosa-fosfato, que mediante el consumo de otra molécula de ATP regenera la ribulosa bisfosfato (RuBP) original, dejándola disponible para que el ciclo se repita nuevamente.
Por tanto, por cada vuelta del ciclo se incorpora una molécula de carbono fijado (CO2) a otra molécula preexistente de 5 átomos de carbono (ribulosa bisfosfato), el resultado final es la regeneración de la molécula de 5 átomos de carbono y la incorporación de un nuevo carbono en forma orgánica C(H2O). Para comprenderlo hay que tener en cuenta que el producto fundamental del ciclo de Calvin es el gliceraldehido-3-fosfato (de 3 átomos de carbono), molécula que sirve como base para la síntesis del resto de carbohidratos. Tras 3 vueltas del ciclo, una nueva molécula de PGAL sale de éste y puede ser posteriormente utilizada para la formación de otras moléculas.
El destino del fosfoglicerato es variado:
a) la mayor parte de él se recicla para regenerar la pentosa fosfato que da inicio al proceso,
b) sintetizar moléculas de glucosa fosfato (vía de las hexosas), ácidos grasos, amoinoácidos... etc; y en general todas las moléculas que necesita la célula. Como por ejemplo dar lugar a la síntesis de almidón, que se conserva en el cloroplasto .
c) abandonar el cloroplasto y dar lugar a la formación de sacarosa o degradarse mediante la vía glicolítica con producción de energía.
Formas alternativas de asimilación del CO2 en plantas
A bajas concentraciones de CO2 (como cuando se cierran los estomas para evitar pérdida de agua en la planta), la Rubisco reaccionara con O2 en vez de CO2. Esta reacción provoca una disminución del porcentaje de carbono fijado y está asociada al fenómeno denominado fotorrespiracion (ver mas adelante). Estos procesos son más graves a temperaturas relativamente altas, disminuyendo la tasa de fotosíntesis (una medida de la capacidad de la planta para asimilar CO2).
Por ello plantas adaptadas a climas cálidos han desarrollado estrategia para optimizar la capacidad de asimilación de dióxido de carbono:
Plantas CAM: Metabolismo Acido de las Crasulaceas:
Es un tipo de metabolismo que se da en plantas y que se descubrió en la familia de las Crassulaceae, de ahí su nombre. El nombre de metabolismo ácido hace referencia a la acumulación de acidos organicos durante la noche por las plantas que poseen este mecanismo de fijación de carbono. Esta vía metabólica es semejante a la vía C4, sin embargo en la vía CAM la separación de los dos carboxilaciones no es espacial, como ocurre en las plantas C4, sino temporal.
Estas plantas tienen dos carboxilaciones separadas temporalmente:
•Fijación nocturna de CO2. Esta primera fase se da en la noche (via de 4 carbonos), cuando tienen los estomas abiertos. A través de ellos la planta capta CO2 atmosférico y la fosfoenolpiruvato carboxilasa lo incorpora por carboxilación al fosfoenolpiruvato (PEP) que se transforma en oxaloacetato (OAA) con el desprendimiento de un grupo fosfato; el oxaloacetato formado de la prefijación de CO2 es reducido en el citosol a malato mediante la NAD-malato deshidrogenasa, el malato es bombeado con gasto de energía a las vacuolas, donde se va acumulando como acido malico y es almacenado, provocando que el contenido vacuolar sea muy ácido (cerca de ph 3) durante la noche.
•Con la salida del sol, los estomas se cierran previniendo la pérdida de agua e impidiendo la adquisición de CO2. El ácido málico sale de la vacuola y se descarboxila liberando el CO2 y acido piruvico el cual es devuelto al ciclo tras ser fosforilado con ATP, produciendo nuevamente nuevamente fosfoenolpiruvato. Ya que los estomas están cerrados, el CO2 liberado internamente no puede escapar de la hoja y en lugar de esto es reducido a carbohidrato por la operación del ciclo C3 PCR. La concentración elevada en el interior de CO2 suprime efectivamente la oxigenación fotorrespiratoria de la ribulosa 1,5-bifosfato y favorece la carboxilación.
Este mecanismo de concentración de dioxido de carbono permite disminuir la probabilidad de que entre un O2 en el sitio activo de la RuBisCO por lo que la eficiencia fotosintética es mayor. Las plantas CAM suelen ser crasas (no todas) y relegadas a ambientes secos (también existen CAM acuáticas); esto es debido a su bajo rendimiento total fotosintético (ya que la absorción de dióxido de carbono está limitado a la cantidad de MA que se puede almacenar en la vacuola) por lo que son malas competidoras con las plantas C3 o C4. Existen plantas CAM constitutivas o adaptativas (estas últimas sólo tienen metabolismo ácido de crasuláceas bajo estrés hídrico, etc.).
Estas plantas resuelven el problema de pérdida de agua durante la fotosíntesis al abrir sus estomas solo durante la noche cuando la temperatura es menor y la humedad del ambiente es comparativamente alta. De manera que el mecanismo CAM le permite a la planta maximizar la eficiencia en el uso de agua. Típicamente una planta CAM pierde de 50 a 100 gramos de agua por cada gramo de CO2 ganado, comparado con los 250 a 300 gramos de la C4 y los 400 a 500 gramos de la C3. Por lo tanto las CAM tienen una ventaja competitiva en ambientes con poca agua, comúnmente se asocian a climas desérticos, pero incluso en ambientes tan húmedos como el bosque tropical es posible encontrarlas en forma de epifitas tales como las orquideas, dado que la cantidad de agua sobre los troncos de sus huéspedes es menor a la registrada sobre el suelo.
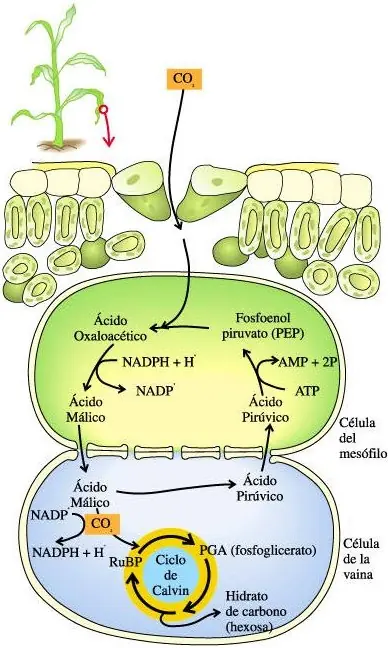
Plantas C-4: Via de 4 Carbonos:
es una serie de reacciones bioquímicas de fijación del carbono proveniente del CO2 atmosférico. El proceso consiste en la captación del Dióxido de carbono en las células del mesofilo de la planta, pero en vez de llevarlo inmediatamente al Ciclo de Calvin, las moléculas reaccionan con el PEP (Fosfoenolpiruvato), siendo estimulados por una enzima homónima (PEP Carboxilasa). El producto final entre el PEP y el CO2 es el ácido oxalacético, el cual posteriormente es convertido en acido malico, o también llamado malato. El malato es llevado a las células de la vaina, en donde es descarboxilado, produciendo el CO2 necesario para el ciclo de Calvin, además de ácido pirúvico. Este último es enviado nuevamente al mesófilo en donde es transformado por medio de ATP en fosfoenolpiruvato (PEP), para quedar nuevamente disponible para el ciclo.
La ventaja de este proceso radica en el hecho de que al tener a la RubisCO encerrada en las células de la vaina se le impide la posibilidad de que reaccione con Oxígeno en situaciones en las cuales la concentración de CO2 sea muy baja, por lo cual el CO2 perdido a través de la fotorrespiracion se reduce considerablemente. Incluso las moléculas de dióxido de carbono expulsadas por la fotorespiración son nuevamente reutilizadas a través del PEP, el cual lo captura en el mesófilo para ser ingresados al ciclo de Calvin.
Las plantas que usan esta vía para la fijación del carbono son denominadas C4, entre las cuales se distinguen el maiz, la caña de azucar, la invasora Cynodon dactylon (Bermuda grass), el sorgo y el amaranto.
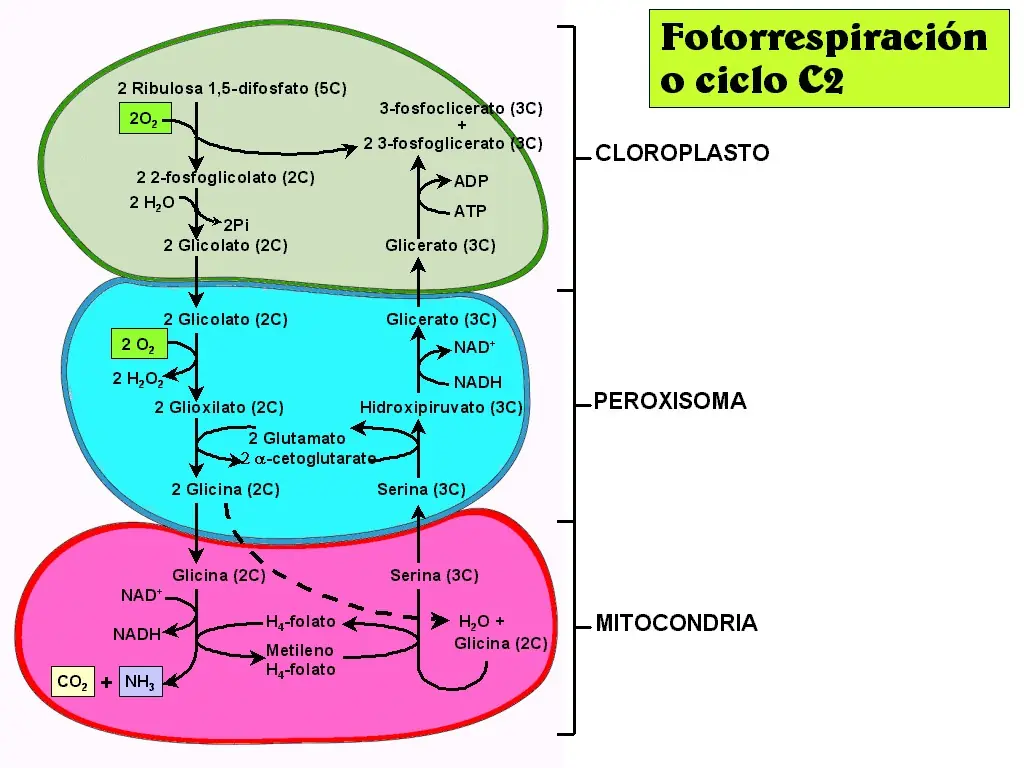
Fotorrespiracion
Una de las propiedades más interesantes de la rubisco es que además de catalizar la carboxilación de la ribulosa 1,5 bifosfato, también produce su oxigenación; proceso conocido como fotorrespiración.
RUBP + O2 + RubisCO ===> Acido fosfoglicerico + Glicolato
La fotorrespiración da como resultado la liberación de CO2 , después de una serie de reacciones enzimáticas. La rubisco de bacterias anaeróbicas autótrofas, cataliza la reacción de la oxigenasa. La reacción de la carboxilación es favorecida a la oxigenación en una proporción de 3:1; lo que indica un 33% de ineficiencia en la carboxilación. El metabolismo del glicolato requiere la participación de las mitocondrias y de los peroxisomas. Sin embargo, es en las mitocondrias donde el aminoácido glicina, producido en los peroxisomas es descarboxilado liberando CO2 .
El ritmo de la fotorrespiración de las plantas C-3 es bastante elevado, siendo 5 veces superior al de la respiración en la oscuridad; lo cual es perjudicial para estas plantas. Las plantas C-4, que muestran muy poca o ninguna fotorrespiración, son considerablemente más eficientes; ya que realizan la fotosíntesis a concentraciones más bajas de CO2 y a más elevadas tensiones de oxígeno.
Las plantas C-4 son de origen principalmente tropical, habitan en condiciones de alta luminosidad y altas temperaturas. Esto les permite competir más eficientemente con las plantas C-3, al tener que cerrar los estomas para economizar agua y evitar la desecación; sin embargo pueden realizar la fotosíntesis a bajas tensiones de CO2 , debido a que la enzima PEP-carboxilasa muestra una mayor afinidad por el CO2 que la rubisco.
La fotorrespiracion es un proceso en el que el cloroplasto absorbe O2, que es catalizado junto con la RuDP por la enzima RubisCO; transformándola así en Ácido Glicólico o Glicolato. El glicolato es traspasado al peroxisoma (saco membranoso que contiene enzimas) y con la acción de O2, son catalizados por la enzima Oxidasa, transformando por una parte en Peróxido de Hidrógeno (Agua oxigenada) y en Glioxilato, el que incorpora nitrogeno por transaminación y forma el aminoácido Glicina. Dos de estos aminoácidos son llevados a la mitocondria donde finalmente se logran tres compuestos: Serina, Amoníaco y CO2. Los gases CO2 y amoniaco se liberan. La serina regresa al peroxisoma en donde es transformada en glicerato, éste es llevado al cloroplasto en dónde, mediante el gasto de una molécula de ATP,se reintegra al ciclo de Calvin como 3-fosfoglicerato.
En conclusión la fotorespiración produce gasto de RuD P y CO2; es un proceso de gasto energético.
Efecto de los herbicidas en el transporte fotosintetico de los electrones
Algunos derivados de la urea, como el monurón o CMU (3-p-clorofenil-1,1 di metil urea) y el DCMU [3-(3,4 di cloro fenil) –1,1 dimetil urea] , se aplican al suelo y se desplazan por el xilema hasta las hoja, donde bloquean el transporte de electrones entre las plastoquinonas QA y QB . Inhiben la reacción de Hill en el fotosistema II¸ por lo que no ocurre la fotólisis del agua ni la liberación de O2 .
Ciertos herbicidas a base de triazinas, como la simazina y atrazina bloquean el transporte de electrones entre QA y QB .El maíz y el sorgo son tolerantes a las triazinas, pero no así a los derivados de la urea; ya que contienen enzimas que detoxifican dichos compuestos. Los herbicidas como el diquat y paraquat ( gramoxone), actúan inhibiendo el flujo de electrones entre la ferrodoxina y el NADP, y reduce el oxígeno a un radical superoxido (O2- ), que produce la perdida de la actividad de los cloroplastos.
CONCLUSION
La fotosíntesis aunque es un proceso bioquímico a nivel celular lo que podría resultar insignificante, tiene mas repercusiones en nuestra vida de lo que podríamos imaginar. Tan solo de ella depende la calidad del aire que respiramos y gracias a ella la atmósfera primitiva se modifico permitiendo así la vida de animales, plantas y el hombre.
La energía solar capturada por el proceso de la fotosíntesis es la fuente de cerca del 90% de toda la energía empleada por el hombre para satisfacer las demandas de calor, de luz y de potencia.
En ella el impacto de los fotones de luz sobre la clorofila y la fotolisis del agua son el origen de un estado de desequilibrio molecular, que se reequilibra constantemente gracias al flujo de protones a través de la membrana de los tilacoides; esto se conoce como Fase Luminosa.
La Fase Oscura consiste en la transformación de dióxido de carbono en glucosa y otros carbohidratos, utilizando para ello la energía química de los productos de la fosforilación. Esta energía almacenada en forma de ATP y NADPH2 se usa para reducir el dióxido de carbono a carbono orgánico. Esta función se lleva a cabo mediante una serie de reacciones llamada ciclo de Calvin, activadas por la energía de ATP y NADPH2. Dando como resultado el 02 liberado a la atmósfera y glucosa que sirve de alimento para la planta.
Cada año, las plantas de nuestro planeta utilizan alrededor de 310,000 millones de toneladas de agua y 750,000 millones de toneladas de dióxido de carbono, para transformarlo en unos 510,000 millones de toneladas de materia y unos 550,000 millones de toneladas de oxígeno.
Suena ilógico que a pesar de que el oxígeno de nuestra atmósfera no se acaba gracias al trabajo de todas las plantas, algas y bacterias, los humanos “supuestamente seres racionales” seguimos talando árboles, contaminando el ambiente, con el smog de automóviles, basura en las calles, aerosoles, humo, etc. Y acabando con todo este oxígeno que es vital para nuestra vida.
BIBLIOGRAFIA
http://es.encarta.msn.com/encyclopedia_761572911/Fotos%C3%ADntesis.html
http://es.wikipedia.org/wiki/Fotos%C3%ADntesis
http://www.forest.ula.ve/~rubenhg/fotosintesis/
http://www.alaquairum.net/fotosintesis.htm
http://www.biologia.edu.ar/plantas/fotosint.htm
http://www.botanica.cnba.uba.ar/Pakete/6to/Fotosintesis/Fotosintesis6to.htm
http://www.educa.aragob.es/iescarin/depart/biogeo/varios/BiologiaCurtis/Seccion%202/2%20-%20Capitulo%209.htm
MATHEWS Y VAN HOLDE. Bioquímica. Edit. McGraw Hill- Interamericana, 2a. edición, 2001.
Hola amigos este es un trabajo sobre fotosintesis el cual hice para la materia quimica biologica, me costo trabajo hacerlo asi q no quiero que otra persona pase por lo mismo,,, espero que les sirva!