Reyben1112
Usuario (Argentina)
Cronologia de una aventura del hombre... Aqui les dejo una pequeña reseña de los mas remotos descubrimientos que dieron origen a la television. Mas alla de todas las criticas que puede tener, la TV cumple un importnte papel como comunicador en el mundo moderno. 1038 Alhazen hace la descripcion de un trompo pintado de diversos colores que permite realizar experiencias sobre el tiempo durante el cual persisten las imagenes en el fondo del ojo. 1039 Alhazen descubre la camara oscura. Es un aparato optico en el que los objetos exteriores se reproducen en el fondo de una caja oscura. La camara oscura en la izquierda y el ojo a la derecha, su similitud es mostrada en esta ilustracion. 1568 Daniel Barbaro recomienda la adicion de un diagrama para hacer más nitida la imagen, controlando la apertura y la cantidad de luz que pasaba a través del lente. Vendrian a representar a nuestras pupilas 1660 El matematico dinamarques Walgestein presenta en Roma una linterna de proyecciones en la cual la luz artificial sustituya la luz solar (aplica la camara oscura a la linterna magica). Primitivo tipo de proyector de imágenes con el que saboyanos y auverneses anónimos asombraron a reyes y gentes de toda Europa desde comienzos del siglo XVII y hasta finales de la década de 1860. 1757 I. B. Beccari descubre el efecto provocado por la accion de luz sobre una placa sensible (cloruro de plata) y aporta un elemento desicivo para la elaboracion de la fotografia. 1802 T. Wedgwood, en un informe presentado a la Royal Institution de Londres, define la fotografia como el medio para fijar imagenes mediante una sustancia sensible a la accion de la luzcon la ayuda de la camara oscura. 1816 Joseph Nicephore Niepce aprovecha los avances quimicos y opticos de los siglos XVII y XVIII, aplica con exito la camara oscura e inventa efectivamente la fotografia. Fija la primera imagen: el tiempo de exposicion empleado es de seis a ocho horas a pleno sol. Era una pequeña imagen negativa en papel emulsionado con cloruro de plata, que se desvanecía al observarla a la luz sin un fijador que la protegiese. 1831 Michael Faraday establece los nexos entre el magnetismo y la electronica y formula las leyes de la induccion electronica. Ley de Faraday establece que el voltaje inducido en un circuito cerrado es directamente proporcional a la rapidez con que cambia en el tiempo el flujo magnético que atraviesa una superficie cualquiera con el circuito como borde 1834 Hoerner, sobre ideas del fisico Plateau, llama zootropio a un aparato de forma cilindrica que gira en torno a un eje vertical. Sobre una cinta de papel colocada en su interior se dibujaban las figuras que representaban sucesivas faces del movimiento. 1839 Louis-Jacques Daguerre revoluciona el mundo de la fotografia al imprimir los primeros daguerrotipos: reduce el tiempo de exposicion a tres o cuatro minutos. Un daguerrotipo es una placa de cobre con un recubrimiento de plata pulida sobre la que se encuentra una imagen de gran detalle y nitidez. Son piezas únicas e irrepetibles ya que la placa se introduce en la cámara obteniendo un solo positivo directo. 1852 Jules Duboscq reemplaza los dibujos de la cinta de papel del zootropio por fotografias. El Bioscope de Jules Duboscq (1850) o stéréofantascope , consistía en dos tambores que giraban de forma sincrónica con los objetivos de un estereoscopio para reconstruir ambos: el relieve y el movimiento. Este sistema podía animar fotografías, pero no podía hacerlas. 1870 Heyl proyecta imagenes fotograficas animadas Un aparato llamado phantasmatrope que consistía en la proyección de imágenes fotográficas móviles. 1871 R. L Maddox introduce el uso de geletina bromuro, con lo cual la exposicion para fijar la imagen se reduce a un centesimo de segundo. 1875 Williams Crooke descubre en Inglaterra los rayos catodicos que se producen en el interior de un tubo de vidrio al vacio. Aqui les dejo una breve explicacion de lo que es: http://es.wikipedia.org/wiki/Tubo_de_Crookes 1877 Emile Reynaud inicia la construccion de un zootropo perfeccionado. Hace proyecciones ante unos 500 espectadores con acompañamientos musicales. Recreación de Émile Reynaud proyectando Pauvre Pierrot. Aplicación óptica desarrollada por Émile Reynaud y patentada en 1888, basada en el praxinoscopio que permite ofrecer un espectáculo a partir de la proyección de dibujos animados móviles, dispuestos en una banda flexible provista de perforaciones que discurren con una frecuencia de 15 imágenes por segundo frente a un sistema de lentes e iluminación basado en la linterna mágica. Es uno de los precedentes del cinematógrafo, sólo que las imágenes habían de dibujarse a mano sobre la película. 1882 J. E. Marey inventa el fusil fotografico, aparato que le permite analizar el movimiento. Demuestra que para descomponer el movimiento se requieren 100 imagenes por segundo. Para mas informacion del fusil , aqui les dejo esto: http://zeodryen.wordpress.com/2008/05/15/historia-del-cine-fusil-fotografico/ 1884 Paul Nipkow patenta un disco giratorio optico-mecanico que permite la transmision de imagenes a distancia. Serviría de mecanismo para proyectar la luz reflejada por un objeto sobre una serie de células de selenio que enviarían los impulsos eléctricos correspondientes a través de un cable. 1886 Heinrich Hertz demuestra la existencia de ondas electromagneticas. Aqui tienen una pequeña bibliografia http://www.biografiasyvidas.com/biografia/h/hertz.htm 1888 George Eastman patenta su camara Kodak. El rollo fotografico es de celuloide. 1896 Guillermino Marconi patenta en Londres la telegrafia sin hilos. Primera proyeccion cinematografica publica realizada por los hermanos Louis y August Lumiere. Louis ha patentado un aparato que sirve obtener proyectar imagenes cronofotograficas. La primera funcion cinematografica se realiza el 22 de marzo de 1895. Se proyecta La sortie des ouvries de l´usine Lumiere. El 28 de diciembre de 1895 un publico curioso pago por el espectaculo cinematografico. link: http://www.youtube.com/watch?v=HI63PUXnVMw 1900 Georges Méliés ensaya la sincronizacion del fonografo con el cine. Aqui les dejo mas info http://es.wikipedia.org/wiki/Georges_M%C3%A9li%C3%A8s 1923 Vladimir Zworykin, fisico estado unidense de origen ruso, inventa el iconoscopio, un analizador elctronico. Las imágenes eran proyectadas en una placa fotosensible, la cual partía la imagen en miles de elementos llamados ahora píxeles. Un rayo de electrones para escaneo, atravesaba la parte frontal de la placa, "cargándola" con todos los píxeles. Cada píxel retenía una carga eléctrica proporcional a la energía luminosa inicialmente proyectada sobre él, el cual era transportado hacia el exterior de la cámara. De esta manera, una imagen visual era convertida en una señal eléctrica. El aspecto clave del Iconoscopio fue que el objetivo integrado era sensible a la imagen, y recolectaba la carga desarrollada por los impactos de luz recibidos entre exploraciones de lectura. 1925 John L. Baird, en Inglaterra, inventa una camara que descompone mecanicamente las imagenes. Se transmiten por cable las primeras imagenes en movimiento. El 26 de enero de 1926, realizó en Londres la primera demostración pública de su sistema ante un grupo de científicos: su muñeco, la vieja marioneta Bill, fue el primer ser en aparecer en una pantalla de televisión. La imagen tenía una resolución de apenas 25 líneas y era diminuta, pero el rostro era perfectamente reconocible. 1929 Zworykin introduce el cinescopio, tubo de rayos catodicos del aparato receptor. El iconoscopio y el cinescopio constituyen los elementos esenciales del sistema television. Tubo de rayos catoticos: El tubo de rayos catódicos (CRT, del inglés Cathode Ray Tube) es una tecnología que permite visualizar imágenes mediante un haz de rayos catódicos constante dirigido contra una pantalla de vidrio recubierta de fósforo y plomo. El fósforo permite reproducir la imagen proveniente del haz de rayos catódicos, mientras que el plomo bloquea los rayos X para proteger al usuario de sus radiaciones. 1930 Primera transmision en vivo: la BBC de Londres al mando de Baird transmiten la tradicional carrera de caballos Derby. 1935 En Inglaterra Marconi-Emi anuncia que producira una imagen con 405 lineas de analisis. 1951 La Columbia emite su primer programa de television en color. 1961 Primera transmision en directo de Moscú a Europa con motivo del regreso del primer vuelo orbital tripulado por Yuri Gagarin. link: http://www.youtube.com/watch?v=CGmphf4CvKw 1962 EE. UU. pone en orbita el Telstar I que funciona como estacion repetidora, lo que hace posible la primera trnsmision de television por satelite entre America y Europa. Se lanzó un segundo Telstar el 7 de mayo de 1963. Su nombre inspiró el del famoso balón de fútbol Adidas Telstar. 1964 El Syncom III permite que las imagenes de los juegos olimpicos de Tokio lleguen a America y Europa. 1965 Se pone en orbita el Intelsat I, primer satelite comercial de comunicaciones, propiedad de los 71 paises miembros del Consorcio Internacional de Comunicaciones por Satelite. Apodado Early Bird o Pájaro madrugador, mas informacion: http://es.wikipedia.org/wiki/Intelsat_I 1969 Primeras imagenes en television en directo desde la Luna, vistas por 723 millones de espectadores. link: http://www.youtube.com/watch?v=QjOxiMNb1oY 1976 Difusion mundial de los juegos olimpicos de Montreal. 1989 Se anuncia que la Argentina esta trabajando en la construccion de un satelite cientifico de comunicacion. La Sociedad Europea de Satelites coloca en el espacio el satelite Astra, que permite recibir en los hogares europeos amplia variedad de canales mediante una pequeña antena parabolica. FUENTE. La informacion aqui publicada fue sacada del libro "Guiones televisivos: antología" Escrito por Eduardo Dayan. Editorial: Ediciones Colihue SRL. Libro incluido en la Coleccion Literarea LYC. Impreso en Bs. As., Argentina, en marzo de 1995. Aca se los dejo por si lo quieren ver: http://books.google.com.ar/books?id=9FmMF8VrphkC&printsec=frontcover&hl=es#v=onepage&q&f=false
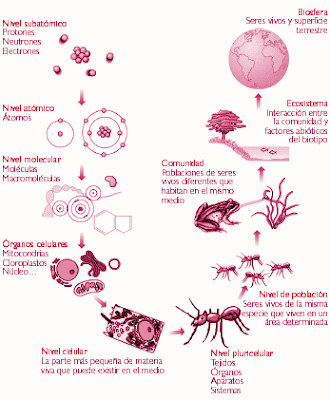
Niveles de organizacion. A los seres vivos se los define por sus características, una de éstas es su organización. Esta organización biológica representa el patrón complejo que nos muestra el camino que ha seguido la evolución, desde formas sencillas a otras más complejas. Los seres vivos están formados por materia. La materia está formada por elementos (92 elementos naturales, como el Cloro, por ejemplo) y se caracteriza por poseer determinadas propiedades intensivas, tales como el punto de fusión, punto de ebullición, conductividad eléctrica, etc. Los elementos están formados por átomos. Un átomo es la porción más pequeña de un elemento que conserva sus propiedades químicas. Las investigaciones de los físicos han descubierto un variado número de partículas subatómicas (Nivel Subatómico), para nuestros fines mencionaremos sólo tres : protones, neutrones y electrones. Los protones son partículas con carga positiva; los electrones, en cambio, tienen carga negativa y masa muy pequeña; los neutrones son partículas neutras, sin carga, y su masa es casi idéntica a la de los protones; los protones y neutrones forman casi toda la masa de un átomo y se localizan en el núcleo atómico. Si combinamos un protón y un electrón se forma un átomo de Hidrógeno, entidad con propiedades diferentes a las de un protón y un electrón (Nivel Atómico). Si combinamos átomos de Hidrógeno entre sí obtenemos Hidrógeno molecular (H2), que es un gas incoloro; si, en cambio, combinamos el H2 con Oxígeno, otro gas, obtenemos agua, una molécula (Nivel Molecular) cuyas propiedades todos conocemos y que no son las mismas que las del H2 y el O2 y que también difieren de las propiedades de las partículas subatómicas y de los átomos que éstas forman. La vida surgió a partir de átomos y moléculas. Si combinamos moléculas entre sí, formamos grandes y complejas moléculas: las macromoléculas, como las proteínas y los ácidos nucleicos (Nivel Macromolecular). Estas macromoléculas constituyen la materia prima que forman los virus (Nivel Prebiótico o Supramolecular) y las células (Nivel Celular). En el Nivel Subcelular múltiples moléculas se ensamblan y dan lugar a estructuras especializadas como los organoides (mitocondrias, cloroplastos, etc). Podemos decir que la vida aparece como propiedad definitoria en el Nivel Celular, o de otro modo, la célula es la porción más sencilla de la materia viva que es capaz de realizar todas las funciones imprescindibles para la vida. En la mayor parte de los individuos pluricelulares, las células se organizan de acuerdo a sus características y funciones conformando tejidos como el conectivo, muscular, epitelial, nervioso (Nivel Tisular). Los tejidos están ordenados en estructuras funcionales, denominadas órganos como el corazón y los pulmones en los animales, o las hojas y las raíces en las plantas. Las funciones biológicas básicas se llevan a cabo por un sistema o aparato, que es una asociación coordinada de tejidos y órganos. Los organismos o individuos pluricelulares están formados por sistemas que actúan en forma coordinada y rigurosa. Existen otros niveles de organización biológica, además de los nombrados anteriormente, donde las propiedades provienen de la relación entre los organismos. Por ejemplo, el Nivel de organización POBLACIÓN reúne a todos los individuos de una misma especie que viven en un mismo lugar, en el mismo tiempo, y que comparten el mismo hábitat. Estas poblaciones interactúan de distinta manera con otras poblaciones del lugar constituyendo una COMUNIDAD, por ejemplo la población de seres humanos de la ciudad de Buenos Aires y el conurbano, aprovecha para alimentarse a las distintas poblaciones de animales y plantas de la zona y se halla parasitada por las mismas poblaciones de parásitos intestinales. Esta comunidad comparte el mismo lugar físico que presenta características particulares. La unión de estos factores físicos con los factores biológicos constituyen los ECOSISTEMAS. Todos los ecosistemas de la Tierra están relacionados, directa o indirectamente. Es por ello que un cambio drástico o continuo de alguno de ellos indefectiblemente acarreará cambios en los restantes. Del mantenimiento de un equilibrio entre los distintos ecosistemas, depende la vida en el planeta. Para terminar de entender les dejo un video explicando este tema... link: http://www.youtube.com/watch?v=DASfZHmcYBI&feature=related Organización Celular Teoría Celular La célula es la unidad de vida más pequeña. Es la unidad anatómica y fisiológica de todos los seres vivos. Dos científicos alemanes el botánico Mattias Schleiden (1804-1881) y el zoólogo Theodor Schwann (1810-1882) fueron los primeros en señalar que "Los cuerpos de las plantas y de los animales están compuestos por células y por productos celulares" enunciando el postulado inicial de la Teoría Celular. Posteriormente, Rudolph Virchow (1821-1902) amplio la Teoría Celular y afirmó: "Todas las células proceden de otra preexistente". Por lo tanto, las células no surgen por generación espontánea a partir de materia inanimada. Posteriormente, Rudolph Virchow (1821-1902) amplio la Teoría Celular y afirmó: "Todas las células proceden de otra preexistente". Por lo tanto, las células no surgen por generación espontánea a partir de materia inanimada. POSTULADOS DE LA TEORIA CELULAR... 1- Todos los seres vivos están formados por células y productos celulares (unidad anatómica) 2- Las funciones de un ser vivo son el resultado de la interacción de las células que lo componen (unidad fisiológica) 3- Toda célula sólo puede tener origen en una célula progenitora. 4- Toda célula tiene la información hereditaria de el organismo del cual forma parte, y esta información pasa de una célula progenitora a una célula hija. Caracteristicas de la celula... Todas las células están cubiertas por una membrana externa, llamada membrana plasmática, que las separa de otras células y del medio circundante con el cual intercambian materia y energía. Este intercambio esta altamente regulado y es selectivo. De esta forma la membrana plasmática debe actuar no sólo como limite celular sino también como barrera selectiva. Por lo tanto la célula, mantiene una composición química muy ordenada y diferente a la del entorno. Todas las células poseen un metabolismo o conjunto de reacciones químicas, que posibilitan el mantenimiento de la vida. Este metabolismo para sustentarse necesita de una o más fuentes de energía. Las células, necesitan de distintivos tipos de moléculas energéticas: * Monedas energéticas, como el ATP * Moléculas combustibles, como la glucosa o los ácidos grasos * Moléculas de reserva de energía, como el glucógeno o el almidón Dentro de las reacciones para obtener e interconvertir diferentes forma de energía, son muy importantes las reacciones de oxido-reducción o reacciones REDOX. En este tipo de reacciones es esencial la participación de las coenzimas de oxido-reducción, como el NAD+ y el FAD. Todas las células, almacenan en forma de ADN, ácido desoxirribonucleico, a información necesaria para controlar sus actividades (reproducción, metabolismo), y para establecer su propia estructura. El ADN, es un polímero formado por una secuencia lineal, de monómeros, llamados nucleótidos. Esta secuencia de nucleótidos, especifica una secuencia de aminoácidos (estructura primaria de una proteína). La especificidad de la secuencia de aminoácidos determinada por la secuencia de bases del ADN esta regida por el código genético. La secuencia de bases del ADN, que codifica una proteína, es un GEN. Las proteínas, son moléculas que llevan a cabo gran parte de las funciones celulares. Muchas proteínas son enzimas, moléculas encargadas de dirigir y regular el metabolismo celular. Las enzimas aceleran las reacciones químicas, haciéndolas compatibles con la vida. De esta manera las enzimas, dirigen la síntesis y degradación de todas las moléculas biológicas, incluidos lípidos, glúcidos, proteínas y los mismos ácidos nucleicos. De esta forma, el ADN al almacenar la estructura de las enzimas y otras proteínas reguladoras, ejerce el control del metabolismo celular. El ADN utiliza un segundo ácido nucleico, el ARN, ácido ribonucleico, como intermediario. A partir de la secuencia de bases del ADN, que codifica una proteína, se sintetiza una secuencia de bases de ARN. Este proceso es llamado transcripción. EL ácido ribonucleico encargado de transportar la información, recibe la denominación de ARN mensajero. Este ARN mensajero, porta la información necesaria para la síntesis de proteínas, proceso llamado traducción, el cual tiene lugar en el citoplasma con la intervención de dicho ARNm, los ribosomas y el ARNt que porta los aminoácidos. Las células para perpetuarse necesitan reproducirse. Esto significa que la información almacenada en el ADN debe duplicarse para poder ser transmitida a las células hijas. El ADN tiene la excepcional característica de ser una molécula capaz de autorreplicarse, es decir de generar una copia de si misma. Este proceso es llamado duplicación o replicación. Dimension Celular... ¿Por qué son tan pequeñas las células? Las células deben captar alimento y otros materiales a través de su membrana plasmática y deben eliminar los productos de desecho, generados en las distintas reacciones metabólicas rápidamente antes de que estos se acumulen hasta niveles tóxicos para la supervivencia celular. Por lo tanto, las células son pequeñas, de modo que en ellas las moléculas recorren distancias cortas, lo que acelera las actividades celulares. Además, a mayor superficie celular, mayor es el transporte de moléculas a través de la membrana, siendo importante para la continuidad de los procesos metabólicos la proporción superficie celular sobre volumen celular. Supongamos una célula de forma cúbica, cuanto más grande es, su superficie crece proporcionalmente lado x lado, es decir a la segunda potencia de la longitud de un lado, en cambio el volumen celular aumenta proporcionalmente a la tercera potencia. Por lo tanto, el volumen celular aumenta más que su superficie a medida que la célula crece, determinando el limite superior al tamaño de la célula en cuestión. Está célula sólo podrá iniciar el proceso de división celular (previa duplicación de su ADN) o perecerá. Por otra parte, debemos recordar que en las células el material Genético (localizado en el núcleo, en células eucariontes), posee un área limitada de influencia sobre el citoplasma circundante, que es el que incrementa marcadamente su tamaño durante el crecimiento celular, siendo otra limitante del tamaño celular la relación núcleo/citoplasma. Como cierre del tema les dejo un videito que me gusto porque da de manera sencilla y entendible una pequeña introduccion al tema... link: http://www.youtube.com/watch?v=9JsLkF4vlmM&feature=related Eso fue todo por ahora, desde ya confio en que les sea util...Espero sus comentarios sobre que les parecio!!! SALUDOS!!!

CARACTERÍSTICAS PRINCIPALES Todas las células se parecen y responden a un patrón común por más diversas que sean. Las células de organismos pluricelulares son diferentes en su función, por ser distintas estructuralmente, pero todas concuerdan con un patrón común. Por ejemplo, aquellas especializadas en la síntesis de lípidos, tendrán mayor desarrollo del retículo endoplasmático liso y serán distintas de las neuronas especializadas en la transmisión del impulso nervioso, cuya especialización es tan grande que pierden su capacidad de reproducirse. A pesar de las semejanzas y diferencias entre las células y que todas cumplen con los postulados de la Teoría Celular, se distinguen dos grandes tipos de células: PROCARIOTAS (sin núcleo verdadero) y EUCARIOTAS (con núcleo). Las características comunes entre los diferentes tipos de células ( animal, vegetal, procariota, eucariota) son: - Todas poseen ADN para la tramsnisión de información bioológica, ARN y Ribosomas para la Síntesis de proteínas por Transcripción y Traducción. - Todas poseen Citoplasma, enzimas metabólicas y membrana plasmática para la Permeabilidad selectiva de tipo activa ( con gasto de energía química ) y pasiva ( sin gasto de energía química). - Todas realizan las mismas actividades metabólicas ( anabolismo-catabolismo). - Todas realizan las 3 funciones fisiológicas, la 1º función de Nutrición en la cual incrporan los nutrientes de otros organismos vivos ( heterótrofa) o bien sintetizan sus propios alimentos por Fotosíntesis ( células Autótrofas), la 2º función de Relación permite a la célula relacionare con el medio externo u otras células, captar estímulos y elaborar una respuesta favorable a ellos ( Sensibilidad o Irritabilidad), la 3º función de Reproducción permite e la célula perpetuarse en el tiempo y transmitir los caracteres hereditarios a la descendencia, puede ser de tipo Asexual de tipo Mitosis ( solo en células eucariotas vegetales y animales de tipo Somáticas o formadoras del cuerpo) asexual de tipo Amitosis o Fisión Binaria( propio de las células procariotas), Sexual o Meiosis ( solo en células eucariotas germinales o Gametas vegetales y animales, Anterozides y Ovocélula en vegetales, óvulo y Espermatozoides en animales). Los eucariontes son organismos cuyas células poseen un sistema de endomembranas (membranas internas) muy desarrollado. Estas membranas internas forman y delimitan organelos donde se llevan a cabo numerosos procesos celulares. De hecho él más sobresaliente de estos organelos es el núcleo, donde se localiza el ADN. Justamente, el término eucarionte, significa núcleo verdadero (eu: verdadero, carion: núcleo). Por lo tanto, las células eucariontes, poseen diversos compartimentos internos, rodeados por membranas. De esta forma es más eficiente reunir a los sustratos y sus enzimas, en una pequeña parte del volumen celular total. Además de conseguirse una mayor velocidad, las membranas favorecen la aparición de estructuras reguladoras que orientan el flujo de moléculas y su posterior conversión en otros productos. Ciertos procesos como la fotosíntesis y la cadena respiratoria están altamente organizados gracias a la localización de las enzimas en diferentes estructuras de membrana. Por otra parte, las membranas también impiden la aparición de sustratos en forma inespecífica en distintas regiones de la célula, ya que actúan como barrera selectiva. En cuanto al tamaño, podemos decir que en promedio una célula eucarionte es diez veces mayor que una célula procarionte. En cuanto al material genético, podemos decir que el ADN eucariota posee una organización mucho más compleja que el ADN procarionte. Las células procariontes carecen de núcleo y generalmente son mucho menores que las células eucariontes. El ADN de las células procariontes no está rodeado por una membrana, pero puede estar limitado a determinadas regiones denominadas nucleoides. Las células procariontes, al igual que las células eucariontes, poseen una membrana plasmática, pero carecen de membranas internas, que formen organelos. Sin embargo, debemos precisar que en algunas células procariontes, la membrana plasmática forma laminillas fotosintéticas. Las células procariontes poseen una característica única, una pared de peptidoglicanos, un gran polímero de glúcidos y aminoácidos. Aca les dejo unos videos que les explican la estructura de las celulas eucariotas y procariotas, de esta ultima se explica la estrucura de una bacteria que es un ejemplo de ese tipo de celula... link: http://www.youtube.com/watch?v=NbYE1f2YYuk&feature=relmfu link: http://www.youtube.com/watch?v=KcFjXYzGh20&feature=relmfu Eso fue todo por ahora, desde ya confio en que les sea util...Espero sus comentarios sobre que les parecio!!! SALUDOS!!!
Hacia fines del siglo pasado se formulo la teoría de que cada enfermedad era producida por un germen específico. Hasta ese momento los patólogos estaban convencidos de que para cada enfermedad seria posible encontrar el microorganismo responsable, utilizando las siguientes técnicas: a) observación del germen con la ayuda del microscopio, b) cultivo sobre un medio nutritivo y c) retención por filtros. Sin embargo, en 1892, Iwanowski (o Ivanovsky?) pudo demostrar que el agente productor de la enfermedad del mosaico de tabaco pasaba a través de los filtros para bacterias y no podía ni verse ni cultivarse. Luego en 1898 Beijerinck, determinó que la enfermedad del mosaico del tabaco era provocada por un nuevo agente infeccioso a los que denomino virus filtrables (virus: palabra de origen latino que significa veneno). Los virus están ampliamente distribuidos en la naturaleza y afectan a todo tipo de organismos, tanto del reino animal, vegetal o protista. Características estructurales de los virus La estructura de los virus esta integrada por dos tipos de macromoléculas: ácidos nucleicos y proteínas, los que forman las partículas virales o viriones. Básicamente existen dos tipos de partículas virales: partículas virales simples (virus desnudo) o partículas virales envueltas (virus envuelto). El virus desnudo consta de un ácido nucleico (genoma) asociado a proteínas y una cubierta proteica o cápside. Por otra parte los virus envueltos añaden a esta estructura básica una envoltura lipoproteica de origen celular. La función de la cápside es de servir al ácido nucleico como protección y vehículo. Postulado de Lwoff "Únicamente serán considerados virus aquellos agentes infecciosos cuya partícula elemental contenga un solo tipo de ácido nucleico". Es decir poseen ARN o ADN, pero no ambos tipos de ácidos nucleicos funcionales a la vez. Por lo tanto los virus pueden ser ADN o ARN virus. Debido a la estructura simple de virus, para su multiplicación dependen en forma absoluta de la célula huésped que infectan. Por lo tanto consideraremos a los virus como parásitos intracelulares obligados. ♠ Replicación viral La célula huésped, una vez infectada, sintetizará nuevas moléculas de ácido nucleico viral, ya que el genoma viral tomara el control de las actividades metabólicas de la célula. Bajo este control, también se producirán gran cantidad de proteínas virales. De esta manera el ensamblado de nuevas partículas virales provendrá de la asociación de las nuevas moléculas de ácido nucleico viral con las proteínas. Este proceso es muy diferente de la reproducción celular, tanto de los procariontes como de los eucariontes. Por lo tanto es mas apropiado hablar de REPLICACION VIRAL. ♠Los virus como parásitos intracelulares Hasta el momento no se ha podido demostrar que ningún virus aislado pueda utilizar o almacenar energía mediante procesos similares a la respiración, tampoco pueden sintetizar proteínas. Es decir no tienen metabolismo propio, dependiendo en forma absoluta del medio ambiente celular. El parasitismo de los virus se ejerce a nivel genético, ya que el genoma viral dentro de la célula desplaza al genoma de la célula hospedadora del control celular. ♠Provirus Anteriormente hemos explicado que los virus, infectan las células y hacen replicas de si mismos. Luego abandonan la célula huésped y pasan a otra, recomenzando su ciclo. Pero también puede ocurrir que el genoma viral se integre al ADN del huésped. Cuando el genoma viral se integra al genoma celular y se replica junto con este se lo denomina PROVIRUS. Un provirus puede activarse espontáneamente o bien expuesto a diversos estímulos, una vez activado puede inducir la producción de virus completos. Los provirus pueden modificar la morfología celular y su metabolismo, esto puede deberse a la producción de alguna proteína viral. Estos cambios en la estructura celular, generalmente asociados a cambios en la membrana celular, inducidos por un provirus reciben el nombre de transformación. En algunos casos estas células transformadas por los provirus pueden ser cancerosas. ♠Los virus como agentes infecciosos El parasitismo celular obligado es la causa básica por la cual un virus puede causar daño. La relación que establece un virus y su célula huésped es variable. El resultado de la interacción depende tanto del virus en cuestión como de la célula huésped. Por ejemplo, se denominan virus citocídicos , a los que como resultado de su multiplicación, producen una rápida inducción hacia la muerte celular. Por otra parte se encuentran los virus no citocídicos, que son aquellos que no provocan la muerte celular. Ejemplos de virus no citocídicos serian los virus moderados y los virus oncogénicos. Los virus moderados son aquellos que producen partículas virales y no producen la muerte celular. Han llegado con la célula huésped a un estado de equilibrio más o menos estable. Luego están los virus oncogénicos, capaces de estimular la división celular, estos cambios pueden ser irreversibles si la célula pierde la capacidad de regular su ciclo celular. Este estado se denomina transformación celular. Los virus no citocídicos pueden causar dos tipos de infecciones: 1- Las infecciones latentes, donde el virus permanece sin manifestarse, alojado en células no productoras. Ante determinados estímulos, estrés, enfermedades, exposición a la luz solar, el virus se reactiva, recomenzando la síntesis de ácidos nucleicos y proteínas virales. Ejemplos: L Herpes simplex, Varicela zoster. 2- Las infecciones crónicas, donde de una persona enferma siempre es posible obtener virus infeccioso, aun por periodos muy prolongados de tiempo. Por ejemplo, virus de la hepatitis B, Epstein-Barr, virus de la rubéola. Morfología viral Los virus poseen gran variedad de formas y tamaños. Por ejemplo los virus que al microscopio electrónico aparecen aproximadamente esféricos se denominan isométricos. En estos virus el ácido nucleico esta rodeado por una cápside (caja) proteica. Las subunidades estructurales que forman la cápside, visibles al microscopio electrónico, se denominan capsómeros. A su vez los capsómeros están formados por subunidades proteicas. La cápside por su naturaleza antigénica es la que determina la identidad viral. El ácido nucleico viral no se encuentra desnudo, sino que esta asociado a proteínas distintas de las de la cápside, cuya función puede ser estructural (plegado del ADN) o enzimática (polimerasa). Al conjunto de ácido nucleico y proteínas asociadas se lo denomina "core". Por ultimo diremos que los virus donde la cápside rodea directamente al ácido nucleico (es decir que no hay un "core" evidente), el conjunto de cápside y ácido nucleico recibe la denominación de nucleocápside. Genoma viral El genoma, que puede encontrase en los distintos tipos de virus, puede sistematizarse de acuerdo a diversos criterios: 1- Tipo de ácido nucleico: ADN o ARN 2- Polaridad o sentido: + o - (aplicado principalmente a los ARN virus) 3- Número de cadenas: monocatenario o bicatenario 4- Genoma circular o desnudo 5- Genoma entero o fragmentado Debemos recordar que las cadenas de un ácido nucleico de doble cadena, son de polaridad (sentido) opuesto. Por convención se considera que si una tiene sentido + la otra será -. De acuerdo a este criterio se considera que un virus es + (o cadena +), cuando su genoma monocatenario tiene la misma polaridad que un ARNm. Es decir el mismo genoma viral, puede actuar dentro de la célula como ARNm y llevar a cabo directamente la síntesis de proteínas virales. Los "virus +", pueden infectar con el ácido nucleico solo, salvo los retrovirus que siendo +, necesitan una transcriptasa reversa asociada al genoma para poder infectar. Por el contrario, se denomina "virus -" cuando el genoma no puede actuar directamente como mensajeros. El ARN - puede actuar como molde, para la síntesis de ARN +, el ARNm viral. Bacteriófagos. Los Bacteriófagos son virus específicos de las bacterias. Bacteriófagos, significa que se "alimenta" o multiplica a expensas de bacterias. Los Bacteriófagos que infectan células huésped, pueden establecer dos tipos de procesos: 1- Ciclo Lítico: en este tipo de ciclo el virus produce inmediatamente gran cantidad de ácidos nucleicos virales y proteínas de la cápside. Estos se ensamblan produciendo nuevas partículas virales que son liberadas al medio al producirse la lisis celular. 2- Ciclo Lisogénico: en este ciclo la relación entre célula huésped y virus, puede prolongase por periodos variables de tiempo. El virus integra su genoma al cromosoma bacteriano, replicándose conjuntamente el ácido nucleico del parásito y el del huésped. Un virus bacteriano integrado al cromosoma se denomina profago. Por lo tanto el profago se replica junto con el ADN bacteriano. En determinadas circunstancias (por ejemplo ruptura del ADN bacteriano por luz ultravioleta o agentes químicos), el profago se activa, y comienza la producción de ácido nucleico viral y proteínas virales, produciendo luego la lisis celular. Las bacterias que portan profagos se denominan lisogénicas. Los Bacteriófagos que pueden integrarse como profagos y que no lisan inmediatamente a las células se denominan fagos atenuados. Les dejo un videito en donde se explica lo de arriba link: http://www.youtube.com/watch?v=KyI8cu-nzRc Estructura Los virus como vectores Los virus pueden servir como vehículos (vectores), para transferir material genético de una célula a otra. Este fenómeno se denomina transducción. Durante el ciclo lítico, el ADN del huésped queda fragmentado y alguno de esos fragmentos puede ser tomado al azar y quedar dentro de la cápside. De esta forma, cuando el virus infecta una nueva célula, transporta genes de una antigua célula huésped a otra nueva. Virus de la Inmunodeficiencia Humana (HIV) El SIDA (síndrome de inmunodeficiencia humana), es una enfermedad infecciosa crónica producida por el virus HIV. Esta enfermedad esta caracterizada por una deficiencia inmunológica progresiva, con la expresión clínica de infecciones oportunistas y/o tumores. El HIV es un retrovirus. Los retrovirus son virus cuyo material genético es ARN. Una vez que el virus ha penetrado en una célula, una enzima llamada retrotranscriptasa, produce ADN a partir del ARN viral. El ADN recién sintetizado viaja al núcleo y se integra al ADN cromosómico . En este punto la infección se ha hecho permanente , y la forma integrada del virus se denomina provirus. El HIV es un LENTIVIRUS. Por lo tanto su ciclo vital intracelular (ciclo de infección) puede prolongarse por años. Sus genomas son complejos y sus mecanismos regulatorios son solo parcialmente conocidos. Los Lentivirus causan infecciones crónicas. El sistema inmune del hombre reconoce las proteínas del HIV como antígenos y produce anticuerpos contra ellas. Por lo tanto una persona infectada tendrá circulando en sangre anticuerpos contra las proteínas del HIV. En este aspecto se basa el test de diagnóstico más utilizado en la actualidad: el test de ELISA (inmunoensayo ligado a enzima). El virión del HIV, esta recubierto por una membrana lipídica, por lo tanto se trata de un virus envuelto. De la membrana sobresalen glicoproteínas: la gp41 y la gp120. La membrana compuesta por lípidos y proteínas recubre el núcleo (core) del virión, formado por las proteínas p28 y p24. En el core se encuentran el ARN del virus y la enzima transcriptasa inversa. Ciclo Vital Intracelular de un retrovirus Empieza cuando un virión o partícula vírica , se une a la superficie externa de una célula susceptible. Este primer estadio del ciclo se lo denomina adsorción. Luego el virus fusiona su envoltura lipoproteica con la membrana celular, introduciendo en la célula su nucleocápside junto con el ARN que constituye su dotación genética. En cada partícula viral se encuentran dos cadenas de ARN vírico. A este proceso se lo conoce como penetración. La enzima transcriptasa inversa es una ADN polimerasa que primero produce una copia de ADN simple cadena que a continuación se copia a si misma obteniéndose ADN doble cadena. Por lo tanto este ADN doble cadena se obtuvo a partir de ARN. La síntesis del ADN doble cadena ocurre junto con la degradación del ARN original. El ADN doble cadena (provirus), emigra hacia el núcleo y se integra en el propio ADN celular. La integración de este ADN doble cadena en el cromosoma del huésped es necesaria para la síntesis de nuevas moléculas de ARN, por la ARN polimerasa celular, ya que el virus carece completamente de la maquinaria metabólica necesaria para realizar la transcripción y la síntesis de proteínas necesarias para la cápside y la misma transcriptasa reversa. Virus de la Hepatitis B (VHB) La hepatitis B en si misma, es un problema sanitario grave y muy extendido . Pero encierra una amenaza peor. El virus que la produce es el carcinógeno humano más importante después del tabaco. Trescientos millones de personas , la mayoría habitantes con escasos recursos asistenciales, están crónicamente infectados con el virus y tienen una probabilidad muy elevada de contraer cáncer de hígado. En el tercer mundo, el virus suele transmitirse de madre a hijo, durante el primer mes de vida, y principalmente durante el nacimiento. Si el pequeño es niña, se convertirá probablemente en portadora crónica y transmitirá el virus de la hepatitis B (VHB) a su descendencia cuando alcance la edad fértil. En cambio en los países desarrollados su incidencia es mayor en adultos, que por su profesión tienen contacto directo con la sangre (cirujanos, enfermeras, dentistas), los receptores de sangre u hemoderivados o personas que reciben tratamientos de diálisis o drogadictos intravenosos. El virus de la hepatitis B (VHB) es mucho más contagioso que el HIV. Viroides. Los viroides son agentes infecciosos que poseen al igual que los virus un solo tipo de ácido nucleico y son parásitos absolutos, pero y esta es la gran diferencia con los virus, carecen de cápside y envoltura. Por lo tanto los viroides están constituidos solo por una secuencia de nucleótidos, además los viroides carecen de información para la síntesis de proteínas, en cambio los virus siempre poseen dicha información. _________________♠♠♠♠_________________ _____________________________________ Priones. Las partículas infecciosas llamadas priones, están constituidas únicamente por una proteína de aproximadamente 250 aminoácidos. Es decir carecen completamente de ácidos nucleicos. Es esta la razón por la cual fue resistida durante mucho tiempo, la hipótesis de que las proteínas por si solas podían ser la causa de enfermedades infecciosas. De acuerdo al dogma imperante hasta 1980, las enfermedades transmisibles (infecciosas) necesitaban material genético, para que la infección se asentara en el huésped. Hasta ese momento eran los virus los agentes infecciosos más pequeños conocidos, y todos ellos poseen ADN o ARN como material genético necesario para codificar sus proteínas y dirigir la replicación viral en el huésped. Pero ahora sabemos que las partículas proteínicas infecciosas (priones), pueden ser el sustrato de diversas enfermedades, hereditarias o contagiosas. Este comportamiento dual tanto infeccioso como hereditario era desconocido. Posteriormente se descubrió que los priones se multiplican por una vía increíble y desconocida hasta ese momento: convierten proteínas normales en MOLECULAS INFECCIOSAS, con solo alterar la estructura proteica. Las encefalopatías espongiformes transmisibles (EET), son las enfermedades degenerativas del sistema nervioso central que afectan a animales y seres humanos causadas por los priones. Se denominan espongiformes ya que el cerebro adquiere un aspecto parecido al de una esponja. Las EET que sufren los seres humanos son el Kuru (o muerte de la risa), la enfermedad de Creutzfeldt -Jakob (ECJ), el síndrome de Gerstman-Straussler-Scheinker (GSS) y el insomnio Familiar Fatal (IFF); las EET de animales, incluyen el scrapie (del ingles to scrape, raspar, por la tendencia de los animales infectados a rasparse contra postes , troncos o cercas para combatir la picazón) de ovejas y cabras, la enfermedad de agotamiento crónico de mulas y ciervos en cautiverio y la encefalitis espongiforme bovina (EEB), o enfermedad de la vaca loca. Las EET se caracterizan por su prolongado periodo de incubación (en el hombre puede tener un periodo de incubación de 30 o mas años), generalmente asociadas a declives progresivos de las funciones motoras y cognitivas (enfermedad activa), y por su evolución inevitablemente fatal. Las EET en el ser humano generalmente aparecen en personas de edad avanzada. Aparentemente todas las EET son causadas por el cambio en la estructura de una proteína normalmente presente en las membranas celulares, denominada proteína priónica (PrP). La forma anormal de la PrP se designa PrPsc (scrapie), para diferenciarla de la forma normal llamada PrPc (celular). La secuencia de aminoácidos (estructura primaria) de la PrPc y la PrPsc es idéntica lo que varia es su conformación (plegamiento en el espacio). De acuerdo a esta teoría la proteína alterada (PrPsc), puede unirse a la proteína normal (PrPc) y cambiar su conformación, transformándola a su vez en una proteína alterada. De esta forma se propagaría la enfermedad y se generarían nuevas proteínas infecciosas. De esta forma el pasaje de la forma normal a la patológica es catalizada por el mismo prión (PrPsc), por lo tanto solo hace falta una pequeña cantidad de este para provocar la transformación de toda la proteína normal, ya que se trata de un fenómeno de crecimiento exponencial. Recientemente , se ha aceptado que los priones pueden ser transmitidos, posiblemente por comida, inoculación directa en el cerebro, piel, músculo o estómago. Por esto la epidemia de EEB, ocurrida en Gran Bretaña provoco un enorme interés en todo el mundo. Desafortunadamente han aparecido en ese país una nueva variedad de la ECJ (casos en personas mucho mas jóvenes que lo usual), lo que probaría una relación causal entre la EEB y los casos seres humanos. El gobierno británico tuvo que admitir la posibilidad de que la aparición de estos extraños casos de la ECJ hubieran sido provocados al ingerirse carne vacuna infectada (tejido nervioso). Al principio habíamos hablado de la dualidad de los priones, por un lado partículas infecciosas y por el otro responsables de enfermedades hereditarias. Esto es naturalmente confuso. Por ejemplo, ciertas enfermedades priónicas como la GSS, son hereditarias. Esta enfermedad tiene una herencia autosomal y dominante, lo cual significa que si un padre desarrolla la enfermedad , los hijos tienen un 50 por ciento de probabilidades de desarrollarla. La explicación a estos hechos vino dado por el descubrimiento de mutaciones génicas puntuales en la secuencia de nucleótidos del gen que codifica la PrP . Estos genes mutados provocan cambios en la secuencia de aminoácidos de la proteína PrP. Estos cambios podrían incrementar la probabilidad de la transformación de la proteína PrP mutante de una forma normal a una anormal patógena. Diferentes mutaciones en el gen provocarían diferentes proteínas mutantes, con mayor o menor tendencia a transformarse en la forma aberrante patógena. Esto explica también las distintas enfermedades priónicas hereditarias, la ECJ esporádica , un 15 por ciento de los casos hereditaria y la GSS autosomal dominante. Eso fue todo por ahora, desde ya confio en que les sea util...Espero sus comentarios sobre que les parecio!!! SALUDOS!!!
Hola a todos!!! Como dicen que andan? Bueno aqui les traigo la segunda parte de mecanica de fluidos. Espero que les sea de ayuda y que comprendan de mejor manera un tema muy lindo como este y que ante cualquier duda sepan consultar sin problemas... La hidrodinamica se ocupa del movimiento de los fluidos en general, y no exclusivamente del movimiento del agua como la raiz etimologica lo pudiera dar a enterder. Definicion de fluido (muy pobre) de la RAE-on line- : http://lema.rae.es/drae/?val=fluido Fluidos segun Wiki: http://es.wikipedia.org/wiki/Fluido El tema que vamos a tratar es propiamente, la dinamica de los fluidos y los conceptos que formulare, mientras no indique otra cosa, son validos en cualquier fluido Solo tendre en cuenta, el movimiento de fluidos en estado estacionario o regimen permanente... Aquel en que las caracteristicas del movimiento no experimentan variacion alguna con el paso de tiempo, es decir, que si en un instante dado por un punto P pasa una particula de fluido con una velocidad V y una aceleracion A, todas las particulas que posteriormente pasen por P lo haran con esa misma velocidad y esa misma aceleracion. (En general para que esto se cumpla en un fluido que pasa por un tubo, el fluido debe tener una velocidad no muy elevada, el tubo tiene paredes lisas y no existan cambios bruscos de seccion). Regimenes de movimiento. El movimiento n regimen estacionario de un liquido en una tuberia puede estudiarse considarando que se trata de un movimiento sin rozamiento (regimen sin rozamiento o de Bernoulli) o de un movimiento laminar (regimen laminar o de Pouseuille). El regimen sin rozamiento corresponde al caso en que la viscosidad del liquido es despreciable. Se caracteriza por el hecho de que las particulas que en un instante dado se encuentran en una misma seccion de la conduccion S1 en cualquier instante posterior siguen estando todas en una misma seccion S2. ¿Que quiere decir esto? Supongamos que tenemos un tubo y en su interior un fluido. En determinada seccion tenemos todo el tubo lleno de determinadas particulas de ese fluido, al ser un liquido que no tiene rozamiento, si lo vemos al mismo liquido en otra seccion, vamos a encontrar esa mismas particulas en la misma posicion unas respecto de las otras como estaban en la primera seccion, es decir ninguna particula se retraso o se adelanto, van todas juntas. El regimen laminar corresponde a un movimiento en regimen estacionario en el que se debe tener en cuenta la viscosidad. En este regimen, las particulas se mueven con mayor velocidad cuanto mas alejadas se encuentran de las paredes de la tuberia, de modo que las que en un mismo instante se encuentran en una misma seccion S1 en un instante posterior ya no lo estaran. ¿Como lo veriamos a esto? Tomemos en cuenta que ahora el tubo, aunque es liso, mientras el fluido pase por él va a quedar mojado, esto quiere decir que al rozar el fluido es como que es frenado por la pared del tubo, esto se debe a la viscosidad. Ahora si vemos en una seccion del tubo unas determinadas particulas, en una segunda seccion vemos que las particulas que estan contra la pared estan frenadas, se retrasan; mientras que las del centro pareciera que se adelantaron. Ecuacion de continuidad. Lo que expresa esta ecuacion es que el caudal que fluye por un tubo es constante, es decir lo que entra va a salir. Considerando un tubo en el que hay un liquido en estado estacionario y dos secciones cuales quieras: Las particulas contenidas en cada una de esas dos secciones habran recorrido una determinada distancia que no sera la misma porque la velocidad no es exactamente la misma en ambos casos. Sean X1= V1. T y X2=V2.T esas distancias. Como los liquidos pueden suponerse incompresibles, los volumenes V1 y V2 barridos por el liquido que pasa por las secciones o areas A1 y A2 deben ser iguales, por lo tanto: Esta expresion nos dice que la velocidad de las particulas de un liquido en los distintos puntos de una conduccion es inversamente proporcional a la seccion. Entonces nos queda que a mas diametro tenga la seccion mas baja es la velocidad del fluido alli y contrariamente a seccion de menor tamaño encontramos mas velocidad. Esto se ve cuando por ejemplo apretamos la punta de una manguera que tira el chorro mas fuerte En resumen: Teorema de Bernoulli. Lo que aqui aparece en juego son las diferencias de presiones y las alturas Este teorema puede ser considerado como la ecuacion fundamental de la hidrodinamica. Constituye una aplicacion del principio de conservacion de energia. Aca les dejo un link en el que explica esto ultimo : https://es.wikipedia.org/wiki/Conservaci%C3%B3n_de_la_energ%C3%ADa Ademas les adjunto un ejemplo de esta ley link: http://www.youtube.com/watch?v=WuEsWnaImLo El teorema de Bernoulli formula la relacion fundamental entre la presion, la altura y la velocidad de un fluido ideal. El mismo demuestra que estas variables no pueden modificarse independientemente una de la otra, sino que estan determinadas por la energia mecanica del sistema. Esta es la ecuacion fundamental de la hidrodinamica, que se reduce a la de la hidrostatica si la velocidad 1 y 2 es 0. Un ejemplo para que entiendan donde se utiliza El funcionamiento de un pulvorizador, ya se trate de uno de tocador o del venturi del carburador de un motor de explosion, se explica facilmente mediante la ecuacion fundamental de la hidrodinamica que hemos visto Vamos a ponerle nombre a cada parte para entender mejor si? Al globito le pondremos A, al tubo que entra en el globo lo llamaremos C, al que viene desde el liquido que se desea pulverizar sera B y el lugar por el que sale despedido puede ser D. Cuando se envia por A una corriente de aire, en C se produce un aumento de la velocidad y, por lo tanto, una disminucion de la presion hidrostatica. El liquido en B asciende hasta C por que tiene menos fuerza que se opone a que suba hasta C y alli se mezcla con el aire, pulverizandose y saliendo por D. Un video en donde explica este tema espero les cierre bien con esto: link: http://www.youtube.com/watch?v=7Cp89RcyYGE link: http://www.youtube.com/watch?v=WI-h4xPsM4w Teorema de Torricelli. El teorema de Torricelli es una aplicación del principio de Bernoulli y estudia el flujo de un líquido contenido en un recipiente, a través de un pequeño orificio, bajo la acción de la gravedad. A partir del teorema de Torricelli se puede calcular el caudal de salida de un líquido por un orificio. "La velocidad de un líquido en una vasija abierta, por un orificio, es la que tendría un cuerpo cualquiera, cayendo libremente en el vacío desde el nivel del líquido hasta el centro de gravedad del orificio": en donde Vt es la velocidad teórica del líquido a la salida del orificio ( cero ), Vo es la velocidad de aproximación o inicial, H es la distancia desde la superficie del líquido al centro del orificio y G es la aceleracion de la gravedad. Para velocidades de aproximación bajas, la mayoría de los casos, la expresión anterior se transforma en: en donde Vr es la velocidad real media del liquido a la salida del orificio y Cv es el coeficiente de velocidad. Para cálculos preliminares en aberturas de pared delgada puede admitirse 0,95 en el caso más desfavorable. Si tomamos Cv= 1 Experimentalmente se ha comprobado que la velocidad media de un chorro de un orificio de pared delgada, es un poco menor que la ideal, debido a la viscosidad del fluido y otros factores tales como la tensión superficial, de ahí el significado de este coeficiente de velocidad. Bueno es todo por hoy, desde ya espero que les haya gustado y que ante cualquier consulta que tengan aca estoy Agradeceria mucho su recomendacion... Pero mas que anda les agradezco el haber pasado Ojala les sea de mucha utilidad. SALUDOS!!!
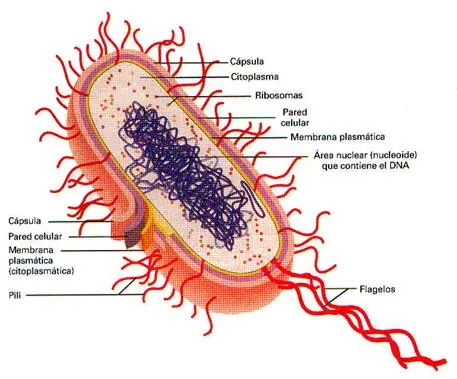
CELULA PROCARIOTA. Las bacterias pueden definirse como organismos unicelulares procariontes que se reproducen por fisión binaria. Contienen toda su información genética en un único cromosoma bacteriano circular. También poseen sistemas productores de energía y biosintéticos necesarios para el crecimiento y la reproducción. Poseen como característica particular una pared rígida de peptidoglicanos. Son generalmente de vida libre y poseen ADN extracromosómico en forma de plásmidos, estos codifican genes de resistencia a antibióticos o factores "sexuales" como los pili. Los micoplasmas son las bacterias mas pequeñas de vida independiente. Son muy flexibles y deformables por lo que atraviesan los filtros de esterilización. Entre sus características principales se encuentran: a) carecen de pared celular, b) en su membrana plasmática poseen esteroles, que no son sintetizados por la bacteria sino que son absorbidos del medio de cultivo o del tejido donde se desarrolla.. Los micoplasmas son resistentes a la penicilina (carecen de pared de peptidoglucano) y por la misma razón no toman la coloración de Gram. Las cianobacterias, anteriormente llamadas algas cianofíceas (azulverdosas), son bacterias Gramnegativas. Se encuentran presentes en estanques, lagos, suelo húmedo, cortezas de árboles, océanos y algunas en fuentes termales. La mayor parte de las cianobacterias son autótrofos fotosintéticos. Contienen clorofila a, que también se encuentra en plantas y algas. La clorofila a y pigmentos accesorios se localizan en membranas fotosintéticas, llamadas laminas internas o laminillas fotosintéticas. Muchas especies de cianobacterias fijan nitrógeno, este proceso enriquece el suelo. ULTRAESTRUCTURA. - Un plásmido es una molécula de ADN extracromosómico que se replica en forma autónoma, por lo que al igual que el cromosoma es un replicón. Puede haber hasta 50 copias de un plásmido en una bacteria. Funcionalmente los plásmidos son elementos genéticos accesorios, es decir que la bacteria puede vivir sin ellos. Sin embargo, la información que contienen puede contribuir a la adaptación de la bacteria al medio y a la evolución de la misma. Los plásmidos pueden contener genes que codifican factores de resistencia a antibióticos (los plásmidos R) y factores de patogenicidad como exotoxinas. La evolución bacteriana a través de los plásmidos es factible, ya que pueden ser intercambiados entre distintas bacterias (por ejemplo, el plásmido F). Es decir que ciertos genes pueden transferirse de una bacteria otra mediante el pasaje de plásmidos, a este mecanismo se lo denomina conjugación. Para que la conjugación pueda llevarse a cabo las dos bacterias deben ponerse en contacto físico . Esto es posible debido a que una de las bacterias, posee pili sexuales (pelos) en su envoltura. Estos pili se encuentran codificados en el mismo plásmido F (plásmido conjugativo). La bacteria que transfiere el plásmido es la que posee pili y se la denomina F+, la célula receptora es F-. - Los transposones son elementos genéticos movibles, que se encuentran presentes en los procariontes (aunque también en las células eucariontes). El descubrimiento de los transposones se lo debemos a Barbara McClintock. Los transposones son fragmentos de ADN que se mueven de una localización a otra del cromosoma. Esta transposición es catalizada por una enzima llamada transposasa. El gen de la transposasa esta incluido dentro del mismo transposón. Los transposones al ser elementos móviles, dentro del genoma, pueden provocar mutaciones al insertarse en nuevas regiones del ADN. - Pared celular. Bacterias Grampositivas y Gramnegativas: Por fuera de la membrana celular, se encuentra una pared celular rígida de peptidoglicano, que esta presente en todas las bacterias excepto los micoplasmas. La presencia de la pared protege a la bacteria de la diferencia de presión osmótica entre el medio interno de la bacteria y el medio exterior. De no existir la pared la bacteria estallaría. Además la pared cumple funciones de protección como por ejemplo contra sustancias tóxicas . Existen dos tipos de pared bacteriana que pueden diferenciarse por la Tinción de Gram (siglo XIX). El primer grupo de bacterias son aquellas capaces de retener el colorante cristal violeta luego de la decoloración con alcohol-cetona. Estas bacterias son llamadas Grampositivas. El segundo grupo esta conformado por aquellas bacterias incapaces de retener el colorante luego del tratamiento decolorante, por lo tanto son llamadas Gramnegativas. Bacterias Grampositivas y Gramnegativas La pared celular de las Grampositivas es más gruesa que la de los Gramnegativas. Posee peptidoglicano, ácidos teicoicos y lipoteicoicos. El componente fundamental es la mureína, un peptidoglicano que solo se encuentra en los procariontes. La mureína consiste en una cadena lineal de dos azúcares alternados N-acetilglucosamina y ácido acetilmurámico. A cada residuo de ácido murámico se encuentra unido un tetrapéptido compuesto de D- y L- aminoácidos. Aproximadamente un tercio de los tetrapéptidos presentes participan de la unión lateral entre cadenas adyacentes de mureína. La pared celular es biológicamente estable, resiste el ataque de las enzimas de los mamíferos, excepto de la lisozima que la degrada. La síntesis de la pared puede ser afectada por antibióticos como la penicilina. Los ácidos teicoicos son el principal determinante antigénico de las bacterias Grampositivas y por lo tanto definen la individualidad inmunológica de estas bacterias. El espesor de la pared celular de una bacteria Gramnegativas es considerablemente menor que el de una Grampositivas. La cantidad de mureína es mucho menor en los Gramnegativas. Los ácidos teicoicos no están presentes en las bacterias Gramnegativas. A ambos lados de la fina pared de mureína se encuentra un gel periplásmico, que define al llamado periplasma (antes llamado espacio periplasmático). Por fuera del periplasma se encuentra una estructura exclusiva de las Gramnegativas, la denominada membrana externa. Si bien es estructuralmente similar a una bicapa lipídica, su composición es diferente de la de otras membranas biológicas. Esta bicapa es muy asimétrica, la semicapa interna esta compuesta por fosfolípidos, pero la semicapa externa esta compuesta por lipopolisacáridos (LPS), altamente tóxico para el ser humano (endotoxina). Para obtener nutrientes las bacterias Gramnegativas, poseen porinas que son proteínas que forman poros en la membrana externa. - Por fuera de la membrana externa de las Gramnegativas y de la gruesa pared de las Grampositivas, se encuentra presente, en algunas bacterias, una cápsula o matriz exopolisacárica, formada por un gel hidrofílico. En general esta cápsula o matriz esta formada por polímeros de azúcares. Las cápsulas permiten a las bacterias evadir los mecanismos de defensa de los organismos pluricelulares, también tienen funciones de adherencia a epitelios permitiendo de esta manera colonizar los tejidos del huésped. Eso fue todo por ahora, desde ya confio en que les sea util...Espero sus comentarios sobre que les parecio!!! SALUDOS!!!
Introducción Las células están separadas del medio que las rodea por una delgada lámina denominada membrana plasmática, que define los límites de las mismas. Hace 3700 millones de años, la formación espontánea de una estructura similar a la membrana plasmática de las células actuales permitió aparición de los primeros seres vivos. Sin esta barrera protectora, las células estarían expuestas a los rigores del mundo externo, no podrían regular su medio interno y, en consecuencia, no serian viables. La membrana plasmática no aísla a la célula completamente sino que constituye una barrera altamente selectiva, que tiene la propiedad de regular el intercambio de materiales entre la célula y el medio que la rodea. La membrana es una estructura muy delgada: sólo tiene un espesor de 6 a 10 nm (1nm=10-9m). Por lo tanto, se necesitarían mil membranas plasmáticas apiladas, una sobre otra, para igualar el espesor de esta hoja de papel. Precisamente debido a su delgadez, cuando se examina una célula al microscopio óptico convencional, puede observarse sin dificultad el interior de la misma; en el mejor de los casos podrá apreciarse el contorno de la membrana, pero nunca podrá distinguirse su ultraestructura. Recién las primeras microfotografías al microscopio electrónico demostraron que la ultraestructura las membranas era siempre la misma. Esta estructura se denominó unidad de membrana y la misma no sólo es válida para la membrana plasmática, sino para casi todas las membranas celulares. Funciones. Como ya se mencionó, las membranas no son simples barreras sino que: · Definen la extensión de la célula y establecen sus límites. · Constituyen barreras selectivamente permeables, dado que impiden el intercambio indiscriminado de sustancias entre el citoplasma y el medio extracelular. La membrana plasmática, gracias a sus propiedades fisicoquímicas, está capacitada para transportar de un lado a otro de la misma determinados solutos, macromoléculas y complejos macromoleculares. Sin embargo, hay moléculas, que a pesar de ser toxicas para la célula, pueden ingresar sin dificultad a la misma a través de la membrana. Un ejemplo seria el CO (monóxido de carbono). · Controlan las interacciones de la célula con el medio extracelular (tanto con la matriz extracelular como con otras células vecinas). Permite a las células reconocerse, adherirse entre sí cuando sea necesario e intercambiar materiales e información. · Intervienen en las respuestas a señales externas a la célula. La membrana posee receptores, que son moléculas o conjuntos de moléculas, capaces de reconocer y responder a señales provenientes del medio extracelular portando información especifica. Cuando dichas señales llegan hasta la membrana plasmática, se desencadenan señales internas en la célula, tanto activadoras como inhibitorias de distintos procesos celulares. Como ejemplos de estas señales externas podemos citar a los factores de crecimiento que favorecen la división celular o diversas hormonas como por ejemplo la insulina, que aumenta la síntesis de glucógeno. Modelo del mosaico fluido. Singer y Nicholson propusieron en 1972 un modelo estructural para las membranas al cual denominaron modelo del mosaico fluido. De acuerdo al mismo las membranas son “disoluciones bidimensionales de lípidos y proteínas.” Según este modelo, la estructura de la membrana sería una delgada lamina formada por dos capas superpuestas de lípidos (también llamadas hemimembranas), con la fluidez propia de los aceites, en la cual se encuentran insertadas proteínas. Esto le confiere el aspecto de un “mosaico”. COMPOSICIÓN Las membranas no son estructuras estáticas ni rígidas. Están formadas por un conjunto de moléculas hidrofóbicas e hidrofílicas que se mantienen unidas por enlaces, en general, no covalentes. Una de las principales características de las membranas biológicas es su alto grado de fluidez. Esto implica que sus lípidos y proteínas pueden desplazarse libremente en todas las direcciones, pero siempre sobre el plano de la membrana. De allí entonces la denominación de “mosaico fluido”; a esta propiedad también se la conoce como difusión lateral. Como puede observarse en el esquema, las membranas también presentan glúcidos unidos por enlaces covalentes a lípidos y proteínas. Esto da lugar a los llamados glucolípidos y glucoproteínas, respectivamente. Estas membranas carecen de resistencia mecánica y en muchas células, como en el caso de hongos, bacterias y plantas están reforzadas por paredes celulares. _____________♦♦♦_____________ _____________________________ Todas las membranas biológicas de los seres vivos, tanto la membrana plasmática, como las de las organelas, están formadas por: Lipidos, Proteinas y Glucidos... La proporción de cada uno de estos componentes varía de acuerdo a la función que realiza cada tipo de membrana. Por ejemplo, las membranas mitocondriales tienen una proporción muy elevada de proteínas. ○Lípidos La variedad de lípidos presentes en las membranas es muy amplia; sin embargo, todos poseen una característica en común: son moléculas anfipáticas. Esto significa que sus moléculas contienen una zona hidrofílica o polar y una hidrofóbica o no polar. Los fosfolípidos son los lípidos más abundantes en las membranas. Debido a su carácter anfipático, los fosfolípidos, en un medio acuoso se organizan espontáneamente conformando la denominada bicapa lipídica. Las cabezas polares están orientadas hacia el medio acuoso (intra y extracelular) y las colas hidrofóbicas hacia el medio lipídico, es decir, al interior de la bicapa, constituyendo la matriz de la membrana. A su vez, estas bicapas tienden a cerrarse espontáneamente sobre sí mismas formando vesículas, es decir, compartimientos cerrados en toda su extensión tridimensional, similares a una esfera. La bicapa de fosfolípidos funciona principalmente como armazón estructural de la membrana y como barrera que impide el pasaje de sustancias hidrosolubles a través de la misma; esto último es debido al carácter fuertemente hidrofóbico de la matriz de la membrana. Los fosfolípidos más frecuentes de las membranas son la fosfatidiletanolamina, la fosfatidilcolina, la fosfatidilserina y la esfingomielina. Los fosfolípidos de las membranas son DIACILGLICERIDOS. La estabilidad de las bicapas lipídicas esta dada por: ♦interacciones hidrofóbicas entre las colas hidrocarbonadas. ♦fuerzas de van der Waals entre las colas hidrofóbicas. ♦fuerzas electrostáticas y puentes hidrogeno entre las cabezas polares de los lípidos, ya sea entre ellos mismos y con las moléculas de agua de los medios extra e intracelular. Como se notará todas estas son uniones débiles (no covalentes) y le confieren simultáneamente estabilidad y fluidez a la membrana. Las cadenas hidrocarbonadas de los ácidos grasos que forman parte los fosfolípidos (también denominadas “colas” o grupos acilo), pueden presentarse: saturados (sin dobles enlaces), monoinsaturados (con un único doble enlace) o poliinsaturados (más de un doble enlace). Ver en la imagen arriba La presencia de ácidos grasos insaturados aumenta la fluidez de la membrana, debido al ”quiebre” de las colas a la altura de los dobles enlaces. Esto impide, o al menos dificulta, que las colas hidrocarbonadas se compacten, restringiendo así las interacciones entre ellas. El hecho de que uno de los grupos acilo de los fosfolípidos esté saturado y el otro no, garantiza una buena fluidez dentro del rango de temperaturas fisiológicas. Por otro lado, cuando las cadenas hidrocarbonadas son cortas, tienen menor superficie para interactuar entre sí; esto último también favorece la fluidez de las membranas. El colesterol es un esteroide que se encuentra en un alto porcentaje en la membrana plasmática de las células animales. Su concentración varía mucho de un tipo de membrana a otro; en animales hay membranas donde el colesterol constituye hasta el 50% del total de los lípidos. Contrariamente, la mayoría de las células vegetales y bacterianas carecen de colesterol. El colesterol, al ser también una molécula anfipática, presenta una orientación similar a la de los fosfolípidos: el grupo hidroxilo (polar) se orienta hacia el exterior de la bicapa y el sector hidrofóbico hacia el interior de la misma. Colesterol en la membrana... Las funciones del colesterol se pueden resumir de la siguiente manera: 1- Inmoviliza los primeros carbonos de las cadenas hidrocarbonadas. Esto hace a la membrana menos deformable y menos fluida, es decir, la estabiliza. Sin colesterol, la membrana necesitaría de una pared celular que le otorgue contención mecánica. 2- Previene el compactamiento de las cadenas hidrocarbonadas a bajas temperaturas, ya que evita que las colas se junten, aumenten las interacciones débiles entre las mismas y se “cristalicen” (adopten una estructura muy compacta). La cardiolipina es un derivado de los fosfolípidos que se encuentra en la membrana interna de la mitocondria. El dolicol es un lípido que se halla en el REG e interviene en la glicosilación de las proteínas. ○Proteinas Mientras que los lípidos ejercen principalmente una función estructural, las proteínas no sólo desempeñan un rol estructural sino que además son las responsables de las funciones específicas de las membranas biológicas. Estas según su función pueden agruparse en: enzimáticas, de transporte, receptoras y de reconocimiento. Diferentes membranas tienen distinta proporción y composición de proteínas, de acuerdo a sus funciones. En otras palabras, son justamente las proteínas las que le otorgan distintas funciones a las membranas. Estas en su mayoría son proteínas globulares (estructura terciaria o cuaternaria). Según su ubicación en la membrana se clasifican en: La imagen esta mostrando la ultraestructura de la membrana, observen la ubicacion de las proteinas: Intrinsecas, Integrales o Transmembrana, atraviesan la membrana como -helice o como láminas plegadas cerradas (estan embebidas en la bicapa). Periféricas o extrinsecas unidas a proteínas transmembrana por interacciones no covalentes débiles y Periféricas unidas a lípidos mediante uniones covalentes (se ven como flotando sobre la membrana). Algunas proteínas multipaso atraviesan muchas veces la membrana y forman un cilindro hueco con un interior hidrofílico por el que pueden pasar moléculas pequeñas solubles en agua. Este es el principio de las proteínas canal que se analizaran mas adelante. Las proteínas integrales pueden difundir lateralmente y rotar sobre su propio eje, pero no pueden realizar movimientos a través del plano de la membrana, o más sencillamente movimiento flip-flop (ver más adelante). Las proteínas integrales suelen desplazarse acompañadas de los lípidos que las rodean ya que estos le ayudan a mantener su conformación. -Proteínas intrínsecas, integrales o transmembrana: Pueden atravesar total o parcialmente la bicapa, asomando a una o ambas superficies de la misma. Únicamente pueden ser extraídas de la membrana por medio de detergentes que rompen la bicapa. Tienen un sector hidrofóbico, que es el que esta insertado en la membrana y una o dos regiones hidrofílicas, expuestas a los medios intra y extracelulares (ambos acuosos). De lo anterior se deduce que estas proteínas son moléculas anfipáticas. La porción que atraviesa la membrana suele presentar una estructura de alfa hélice con una elevada proporción de aminoácidos hidrofóbicos que interaccionan con las colas hidrocarbonadas de la matriz de la membrana. El sector proteico (también llamado dominio) expuesto a los medios acuosos suele tener estructura globular e interacciona con las cabezas polares de los fosfolípidos y con otras moléculas a través de uniones iónicas y puente de hidrógeno. Dentro de las proteínas integrales encontramos: ♦ Proteínas monopaso: La proteína “atraviesa” una sola vez la membrana. ♦ Proteínas multipaso: La cadena polipeptídica atraviesa dos o más veces la bicapa lipídica. Por lo tanto, esta posee varias regiones hidrofóbicas insertadas en la matriz de la membrana alternadas con sectores hidrofílicos que se exponen hacia los medios acuosos. Algunas proteínas multipaso atraviesan muchas veces la membrana y forman un cilindro hueco con un interior hidrofílico por el que pueden pasar moléculas pequeñas solubles en agua. Este es el principio de las proteínas canal que se analizaran mas adelante. Las proteínas integrales pueden difundir lateralmente y rotar sobre su propio eje, pero no pueden realizar movimientos a través del plano de la membrana, o más sencillamente movimiento flip-flop (ver más adelante). Las proteínas integrales suelen desplazarse acompañadas de los lípidos que las rodean ya que estos le ayudan a mantener su conformación. Sin embargo, algunas proteínas integrales están ancladas a componentes del citoesqueleto y no pueden trasladarse. De esta manera intervienen en la morfología de la célula, por ejemplo alargada (o ahusada), cúbica, cilíndrica, etc. -Proteínas extrínsecas o periféricas: Se encuentran sobre la cara externa o también interna de la membrana y pueden estar ligadas tanto a las proteínas integrales como a los fosfolípidos por uniones débiles. Se pueden extraer fácilmente con tratamientos no drásticos. Cuando estas se ubican del lado citoplasmático de la membrana suelen interactuar con el citoesqueleto. ○Hidratos de carbono Las membranas celulares contienen entre un 2-10% de glúcidos. Estos se asocian covalentemente a los lípidos (glicolípidos) y a las proteínas (glicoproteínas). Los glicolípidos (o glucolipidos) presentes en las membranas son los gangliósidos y cerebrósidos. Los gangliósidos se forman por la unión de un oligosacárido con la ceramida. La estructura de los cerebrósidos es similar, sólo que el hidrato de carbono no es un oligosacárido sino una galactosa o una glucosa. Los hidratos de carbono de los glucolípidos y las glucoproteínas, en su mayoría oligosacáridos, suelen ubicarse en la cara no citosólica de la membrana plasmática formando una estructura llamada glicocálix. Las diferencias entre los grupos sanguíneos se hallan determinadas por ciertos oligosacáridos muy cortos, presentes en las membranas plasmáticas de los glóbulos rojos o eritrocitos. Estos oligosacáridos sólo difieren en sus monómeros terminales y están ligados a una proteína transmembranosa o a una ceramida de la membrana plasmática. Por ejemplo, los eritrocitos pertenecientes al grupo sanguíneo A, presentan como monosacárido terminal una N-acetilgalactosamina y los del grupo B una galactosa. Cuando ambos monosacáridos terminales están ausentes estamos en presencia del grupo 0. Propiedades de la membrana... La fluidez de la membrana y los factores que influyen en ella. Como ya se mencionó, las membranas son estructuras dinámicas donde los componentes pueden desplazarse en todas las direcciones sobre el plano de la bicapa. De ahí que el modelo reciba el nombre de mosaico fluido. Existen tres tipos de movimientos posibles en las membranas: ♦ rotación (sobre su propio eje) ♦ traslación (o difusión lateral) sobre el plano de la membrana. ♦ flip-flop El movimiento de flip-flop es el intercambio de fosfolípidos de una monocapa (o hemimembrana) a la otra; esta sumamente restringido, debido a la dificultad que posee la cabeza polar para atravesar el medio hidrofóbico de la matriz de la membrana. De allí que no sea un movimiento que ocurra de manera espontánea sino que está mediado por enzimas denominadas flipasas. Tanto los movimientos de difusión lateral como el de rotación se llevan a cabo sobre la misma hemimembrana de la bicapa lipídica. Factores que aumentan la viscosidad de las membranas - Ácidos grasos insaturados - Baja concentración de colesterol - Altas temperaturas - Colas hidrocarbonadas cortas (dificultan el empaquetamiento) Factores que favorecen la viscosidad: ♦Alto grado de saturación y mayor longitud de las colas hidrocarbonadas. ♦Menor temperatura del medio. Factores que favorecen la fluidez: ♦Alto de grado de insaturación y menor longitud de las colas hidrocarbonadas. ♦Mayor temperatura del medio. El ascenso de la temperatura aumenta la energía cinética entre las moléculas y, por lo tanto, el movimiento de las colas hidrocarbonadas. Esto lleva a una disminución de las interacciones atractivas entre las mismos y a un aumento de los movimientos de rotación y de difusión lateral. Por el contrario, una disminución de la temperatura vuelve más rígida a la membrana ya “empaqueta” las colas hidrofóbicas de los fosfolípidos e impide sus movimientos. Si la temperatura desciende significativamente, la membrana puede llegar a “cristalizarse”, con la pérdida consiguiente de muchas funciones vitales de la membrana. Los organismos que habitan regiones donde hay grandes amplitudes térmicas estacionales varían la composición de los fosfolípidos de sus membranas en forma periódica, asegurando así una fluidez más o menos constante durante todo el año. Por otra parte, organismos que habitan ambientes extremos poseen composiciones fosfolipídicas muy particulares en sus membranas, por ejemplo, los que viven a temperaturas inferiores a los 0ºC tienen membranas muy ricas en lípidos poliinsaturados. La fluidez de la membrana se pudo determinar experimentalmente tratando células con anticuerpos fluorescentes que eran reconocidos y se unían a las proteínas (receptores) presentes en la membrana plasmática. Gracias a esta técnica, se pudo observar, a través del microscopio, el desplazamiento de los receptores sobre la superficie de la membrana y su agrupamiento en un polo de la célula, donde posteriormente ingresaban por endocitosis. La asimetría estructural de las membranas. En ambas caras de la bicapa (también denominadas hemimembranas o monocapas) no se encuentran los mismos tipos de fosfolípidos. Si bien estos en su mayoría se sintetizan en la cara citosolica del retículo endoplasmático liso, luego, por medio de movimientos del tipo flip-flop (únicamente permitidos en el REL, gracias a la presencia de flipasas), se van ubicando del lado de la bicapa que les corresponda. Por ej., la fosfatidilcolina y la esfingomielina predominan en la cara no citosolica de la membrana. La asimetría estructural de las membranas suele manifestarse a través de una asimetría funcional. Esto significa que las funciones presentes en la cara citosolica no son las mismas que aparecen en la cara no citosólica. Por ejemplo, en el caso de la membrana plasmática, las moléculas que intervienen en el reconocimiento celular se ubican casi exclusivamente en la cara expuesta hacia el medio extracelular, pues no tendría mucho sentido que dichas moléculas estuviesen expuestas hacia el citoplasma. Fusion de membranas... Las membranas tienen una elevada capacidad para fusionarse entre sí. Por ejemplo, cuando una vesícula se aproxima a la membrana plasmática, a una cisterna o, inclusive, a otra vesícula, al entrar en contacto ambas superficies, las dos membranas se fusionan, constituyendo a partir de ese momento una sola membrana. Este fenómeno explica el transito de sustancias desde un compartimiento celular a otro, y desde las endomembranas a la membrana plasmática. Este es el principio en el que se basa la administración de fármacos vehiculizadas dentro de liposomas, que son vesículas fosfolipídicas artificiales que contienen alguna droga de interés terapéutico. Cuando el liposoma se aproxima a la célula blanco (o target), la membrana del liposoma se fusiona con la membrana plasmática liberando su contenido directamente en el citoplasma de la célula. Este fenómeno permite que el contenido del liposoma sólo sea captado por ciertos tipos celulares y no por otros. Técnicas basadas en esta propiedad de las membranas se utilizan, por ejemplo, para combatir células tumorales. Eso fue todo por ahora, desde ya confio en que les sea util...Espero sus comentarios sobre que les parecio!!! SALUDOS!!!
Huesos de la Cara Cara: Es un conglomerado óseo, situado en la parte inferior y anterior de la Cabeza. • Contiene en sus cavidades la mayoría de los aparatos de los sentidos. • En la cara hay catorce huesos. • Doce son formados por seis pares. • Los otros dos son impares o únicos y se localizan en la línea media. Maxilar superior: • Es un par de huesos cortos e irregulares. • Aplanado de adentro afuera. • Presenta dos caras interna y externa. • Cuatro bordes y cuatro ángulos. • En su borde inferior da inserción a los dientes de la arcada superior. Se articula con: • El maxilar del lado opuesto en la línea media. • Con el frontal y el etmoides y los huesos propios de la nariz por arriba. • Los palatinos y el vómer hacia el medio y por detrás. Contribuyen en la formación de la órbita ocular y de las fosas nasales. Palatinos: • Son un par de huesos cortos e irregulares. • Ocupan uno en el lado derecho y otro en el izquierdo. •Presentan para su estudio dos láminas, una horizontal y otra vertical. •Se localizan por detrás del maxilar con quien se articula hacia delante. •Se articula con el palatino del lado opuesto. •Por detrás se articula con el esfenoides. • Por arriba con el vómer y conchas nasales inferiores. Contribuye a la formación de las fosas nasales. Cigomaticos: •Hueso par, corto e irregular situado en la parte más externa de la cara. •Aplanado de fuera adentro. •También conocidos como malares o Pómulos. •Forma cuadrilátera. Presenta: •Dos caras externa e interna. • Cuatro bordes. •Cuatro ángulos. Se encuentran en la cara inferior y lateral al frontal. Se articulan: • Por arriba con el frontal. •Por debajo con los maxilares Superiores. •Por los lados con los temporales. Contribuyen a la formación de la órbita ocular. Huesos propios de la nariz o nasales: •Hueso par colocado a cada lado de la línea media. •Es una lámina cuadrilátera con dos caras y cuatro bordes. •Se articulan con: •Por arriba con el frontal. •Por debajo con el maxilar Superior. •Con el homónimo del lado opuesto en la línea media y con el etmoides. Contribuyen con la formación de las fosas nasales. Cornetes o Conchas Nasales Inferiores: •Hueso par, situado en la parte inferior de las fosas nasales. • Se encuentra en la parte inferior de las fosas nasales, la cual contribuyen a formar. •Presentan para su estudio: •Dos caras internas interna y externa. • Dos bordes. • Dos extremidades. Se articulan con: •El etmoides y el maxilar superior por arriba. •Con el palatino por detrás. •Con los lagrimales por delante. Unguis o huesos Lagrimales: Son un par de huesos, que se hayan situados en la parte anterior de la cara interna de la fosa orbitaria. •También contribuyen a formar las fosas nasales. •Constituye una pequeña lámina ósea. •Forma cuadrilátera irregular. •Tiene dos caras y cuatrobordes. Vomer: • Es un hueso único o impar. • Se encuentra en la línea media de la cara. •Constituye la parte posterior del tabique nasal. Lámina cuadrilátera muy delgada. •Presenta para su estudio: •Dos caras. • Dos bordes. Se articula con: •El etmoides y el esfenoides por arriba. •Los maxilares superiores y los palatinos por debajo. Forma parte del tabique Nasal. Maxilar Inferior o Mandíbula: •Hueso grande, único, irregular, simétrico y central. • Se localiza en la parte inferior de la cara. •Tiene forma de herradura. •Único hueso de la cara que se une a otros huesos por una articulación móvil. •Presenta para su estudio: •Dos caras, anterior y posterior. • Dos extremidades laterales o ramas ascendentes. • Dos Bordes. En el superior da inserción a los dientes de la arcada inferior. ______________________________________________♦♦♦♦_________________________________________ ___________________________________________________________________________________________ Ademas les dejo unos videos del Dr Leonardo Coscarelli...que desarrollan mas en profundidad el tema!!! link: http://www.youtube.com/watch?feature=player_embedded&v=r0Anh_2B-MQ link: http://www.youtube.com/watch?feature=player_embedded&v=bZtPC-F3XeA link: http://www.youtube.com/watch?feature=player_embedded&v=T7d2VAqJWPw link: http://www.youtube.com/watch?feature=player_embedded&v=fvtizPaChx0 link: http://www.youtube.com/watch?feature=player_embedded&v=LRcJSu3e8-o link: http://www.youtube.com/watch?feature=player_embedded&v=sIzEak4g0Rk ________________________________________♦♦♦♦_______________________________________ ___________________________________________________________________________________ Bueno eso fue todo!!! Espero les sea de mucha utilidad... SALUDOS...

Introduccion. Como muchos sabrán, para poder tener energía para afrontar las distintas actividades que realizamos día a día es necesario alimentarnos debidamente, incorporando Vitaminas y Proteínas, y contando con el ingreso de nuestra principal fuente energética como lo es la Glucosa, que está presente en una alta riqueza en la naturaleza, siendo la base de los productos de Almidón que preparan las plantas y organismos fotosintéticos a base de Agua y Nutrientes del Suelo. También otro de los derivados de la Glucosa está presente en la Celulosa, como también los azúcares que encontramos en Vegetales y Frutas, o también otros productos que se elaboran en base a ellos, como la Fructosa y la Sacarosa, que ingresan en nuestro cuerpo mediante los alimentos y en distintas formas químicas, que luego se transforman en otras mediante distintos Proceso Metabólicos. El conjunto de estas operaciones que realiza nuestro cuerpo automáticamente lleva el nombre de Glucólisis, y es básicamente la forma en la que nuestro cuerpo parte de la forma de una Sustancia Compleja derivada de la glucosa, hacia la obtención de una sustancia que pueda ser aprovechada por nuestro organismo, para la obtención de energía. Nuestros músculos obtienen la energía necesaria para poder movilizarse y realizar distintas actividades a través de Moléculas de ATP, que son obtenidas mediante la ruptura de moléculas de Glucosa (es decir, una Lisis de Glucosa, de allí el término) lo que permite no solo brindar una alimentación a todo el cuerpo, sino fundamentalmente a Nivel Celular, permitiendo el desarrollo de nuevas células y un marcado crecimiento. Seguramente muchos hemos oído que antes de hacer una fuerte actividad física necesitamos una ingesta abundante de Hidratos de Carbono, ya que éstos generalmente son distintas fomas del Almidón proveniente de las Frutas y Vegetales, siendo además los que se metabolizan más rápido (lo que es comúnmente llamado “Digestión Rápida”) y cumpliendo a la perfección con las exigencias físicas, sobre todo deportivas. La glucólisis es la principal ruta para el metabolismo de la glucosa y la principal vía para el metabolismo de la fructosa, galactosa y otros carbohidratos derivados de la dieta. La capacidad de la glucólisis para proporcionar ATP en ausencia de oxígeno tiene especial importancia, porque esto permite al músculo esquelético tener un nivel muy alto de desempeño cuando el aporte de oxígeno es insuficiente, y permite a los tejidos sobrevivir a episodios de anoxia. Las enfermedades en las cuales hay deficiencia de las enzimas de la glucólisis (p. ej., piruvato cinasa) se observan sobre todo como anemias hemolíticas o, si el defecto afecta el músculo esquelético (p. ej., fosfofructocinasa), como fatiga. En las células cancerosas en crecimiento rápido, la glucólisis procede a un índice alto, formando grandes cantidades de piruvato, el cual es reducido hacia lactato y exportado. Esto produce un ambiente local hasta cierto punto ácido en el tumor, mismo que puede tener inferencias para la terapia del cáncer. El lactato se usa para gluconeogénesis en el hígado, proceso costoso en cuanto a energía, del cual depende gran parte del hipermetabolismo que se observa en la caquexia por cáncer. La acidosis láctica se produce por varias causas, entre ellas actividad alterada de la piruvato deshidrogenasa. Glucolisis en anaerobiosis. En las primeras investigaciones de la glucólisis quedó de manifiesto que la fermentación en levaduras era similar a la degradación de glucógeno en el músculo. Fue evidente que cuando un músculo se contrae en un medio anaerobio, esto es, uno a partir del cual se excluye el oxígeno, el glucógeno desaparece y aparece lactato; cuando se admite oxígeno, tiene lugar la recuperación aerobia, y ya no se produce lactato. No obstante, si ocurre contracción en condiciones aerobias, no hay acumulación de lactato, y el piruvato es el principal producto terminal de la glucólisis. El piruvato se oxida más hacia CO2 y agua. Cuando hay carencia de oxígeno, la reoxidación mitocondrial de NADH formado durante la glucólisis está disminuida, y el NADH se reoxida al reducir piruvato a lactato, de modo que se permite que proceda la glucólisis. Si bien la glucólisis puede ocurrir en condiciones anaerobias, esto tiene un precio, puesto que limita la cantidad de ATP formado por cada mol de glucosa oxidada, de modo que debe metabolizarse mucho más glucosa en condiciones anaerobias que en condiciones aerobias. En levaduras y algunos otros microorganismos, el piruvato formado en la glucólisis anaerobia no se reduce a lactato, sino que se descarboxila y reduce a etanol. Glucolisis. Todas las enzimas de la glucólisis se encuentran en el citosol. La glucosa entra a la glucólisis por medio de fosforilación hacia glucosa 6-fosfato, catalizada por la hexocinasa, usando ATP como el donador de fosfato. En condiciones fisiológicas, la fosforilación de glucosa hacia glucosa 6-fosfato puede considerarse irreversible. La hexocinasa es inhibida de manera alostérica por su producto, la glucosa 6-fosfato. En tejidos que no son el hígado (y en las células β de los islotes pancreáticos), la disponibilidad de glucosa para glucólisis (o para síntesis de glucógeno en el músculo y lipogénesis en el tejido adiposo) se controla mediante transporte hacia la célula, que a su vez está regulado por la insulina. La hexocinasa tiene afinidad alta (Km baja) por la glucosa, y en el hígado está saturada en condiciones normales y, así, actúa a una velocidad constante para proporcionar glucosa 6-fosfato para satisfacer las necesidades de la célula. Las células del hígado también contienen una isoenzima de la hexocinasa, la glucocinasa, que tiene una Km mucho más alta que la concentración intracelular normal de glucosa. La función de la glucocinasa en el hígado es eliminar glucosa de la sangre después de una comida, proporcionando glucosa 6-fosfato en una cantidad superior a los requerimientos para la glucólisis, que se usa para la síntesis de glucógeno y lipogénesis. La glucosa 6-fosfato es un importante compuesto en la unión de varias vías metabólicas: glucólisis, gluconeogénesis, la vía de la pentosa fosfato, glucogénesis y glucogenólisis. En la glucólisis se convierte en fructosa 6-fosfato mediante la fosfohexosa isomerasa, que comprende una isomerización aldosa-cetosa. Esta reacción va seguida por otra fosforilación catalizada por la enzima fosfofructocinasa (fosfofructocinasa-1) que forma fructosa 1,6-bisfosfato. En condiciones fisiológicas puede considerarse que la reacción de la fosfofructocinasa es funcionalmente irreversible; es inducible, está sujeta a regulación alostérica, y tiene una participación importante en la regulación del índice de glucólisis. La aldolasa (fructosa 1,6-bisfosfato aldolasa) divide a la fructosa 1,6-bisfosfato en dos triosa fosfatos, el gliceraldehído 3-fosfato y dihidroxiacetona fosfato. La enzima fosfotriosa isomerasa interconvierte estos dos últimos compuestos. La glucólisis continúa con la oxidación de gliceraldehído 3-fosfato a 1,3-bisfosfoglicerato. La enzima que cataliza esta oxidación, gliceraldehído 3-fosfato deshidrogenasa, es dependiente del NAD. Desde el punto de vista estructural consta de cuatro polipéptidos idénticos (monómeros) que forman un tetrámero. En cada polipéptido hay cuatro grupos —SH, derivados de residuos cisteína dentro de la cadena polipeptídica. Uno de los grupos —SH se encuentra en el sitio activo de la enzima. El sustrato inicialmente se combina con este grupo —SH, lo que forma un tiohemiacetal que se oxida hacia un tiol éster; los hidrógenos removidos en esta oxidación se transfieren al NAD+. El tiol éster pasa después por fosforólisis; se agrega fosfato inorgánico (Pi), lo que forma 1,3-bisfosfoglicerato, y el grupo —SH se reconstituye. En la reacción siguiente, catalizadapor la fosfoglicerato cinasa, el fosfato se transfiere desde el 1,3-bisfosfoglicerato hacia ADP, lo que forma ATP (fosforilación a nivel de sustrato) y 3-fosfoglicerato. Dado que se forman dos moléculas de triosa fosfato por cada molécula de glucosa que pasa por glucólisis, en esta etapa se forman dos moléculas de ATP por cada molécula de glucosa que pasa por glucólisis (se recuperan las 2 utilizadas en las reacciones anteriores. La toxicidad del arsénico resulta de la competencia del arsenato con el fosfato inorgánico (Pi) en esta reacción anterior para dar 1-arseno-3-fosfoglicerato, que se hidroliza de manera espontánea hacia 3-fosfoglicerato sin formar ATP. La fosfoglicerato mutasa isomeriza el 3-fosfoglicerato hacia 2-fosfoglicerato. Es probable que el 2,3-bisfosfoglicerato (difosfoglicerato, DPG) sea un intermediario en esta reacción. El paso subsiguiente es catalizado por la enolasa, y comprende una deshidratación, lo que forma fosfoenolpiruvato. La enolasa es inhibida por el fluoruro, y cuando se obtienen muestras de sangre para medición de glucosa, se deben recolectar en tubos que contengan fluoruro a fin de inhibir la glucólisis. La enzima también depende de la presencia de Mg2+ o Mn2+. El fosfato del fosfoenolpiruvato se transfiere hacia el ADP mediante la piruvato cinasa para formar dos moléculas de ATP por cada molécula de glucosa oxidada. La reacción catalizada por la piruvato quinasa es otro ejemplo de fosforilación a nivel de sustrato: la conversión del compuesto de alta energía fosfoenolpiruvato se transforma en enolpiruvato y libera energía permitiendo la síntesis de ATP. Esta reacción es también otro punto importante de regulación de la vía glucolítica. Esta enzima es inhibida por ATP lo que permite disminuir la velocidad de la vía glucolítica cuando la carga energética de la célula es alta. La alanina también inhibe su actividad. La fructosa 1,6 di fosfato, producto de la reacción irreversible precedente de la vía glucolítica, es un activador de esta enzima, lo que evita el acumulo de intermediarios de la vía cuando se produce la activación de las enzimas regulatorias de los pasos previos La actividad de la enzima está también regulada por modificación covalente bajo control hormonal. Se han caracterizado tres isoenzimas de la piruvato quinasa en mamíferos: La de tipo L predomina en hígado, la tipo M en músculo y cerebro y la de tipo A, en otros tejidos. La fosforilación de la isoenzima L de la piruvato quinasa por una quinasa dependiente de AMPc, la inactiva. Desfosforilada es activa. Cuando la glucosa sanguínea es baja, se incrementa en sangre el nivel de la hormona glucagon, una hormona que se une a un receptor de la membrana plasmática y produce activación de la adenilato ciclasa y consecuentemente un incremento en los niveles intracelulares de AMPc y activación de la proteína quinasa AMPc dependiente. Esta cascada de eventos desencadenada por acción hormonal conduce a la fosforilación de la isoenzima L, la cual predomina en hígado, y lleva a su inactivación. De manera que una disminución de la glucemia lleva a la inactivación de la vía glucolítica en el hígado, evitando de esta manera el consumo de glucosa por este tejido cuando esta es más necesaria por ejemplo para el cerebro. El estado redox del tejido ahora determina cuál de dos vías se sigue. En condiciones anaerobias, el NADH no se puede reoxidar por medio de la cadena respiratoria a oxígeno. El NADH reduce el piruvato hacia lactato, lo cual es catalizado por la lactato deshidrogenasa. Hay diferentes isoenzimas de lactato deshidrogenasa específicas para tejido, que tienen importancia clínica. La reoxidación de NADH por medio de la formación de lactato permite que la glucólisis proceda en ausencia de oxígeno al regenerar suficiente NAD+ para otro ciclo de la reacción catalizada por gliceraldehído 3-fosfato deshidrogenasa. En condiciones aerobias, el piruvato es captado hacia las mitocondrias, y después de descarboxilación oxidativa hacia acetil- CoA, el ciclo del ácido cítrico lo oxida hacia CO2. Los equivalentes reductores provenientes del NADH formado en la glucólisis son captados hacia las mitocondrias para oxidación por medio de uno de transbordadores (lanzaderas). Regulacion de la glucolisis. La regulación de la vía es ejercida por distintos mecanismos. Fue mencionada la importancia de las enzimas fosfofructoquinasa I y piruvato quinasa en este aspecto. Distintos efectores modulan la actividad de estas enzimas. Por ejemplo, si la carga energética celular se eleva, ATP/ADP, se produce la inhibición de ambas enzimas lo cual determina la disminución de la velocidad de la vía. La acción hormonal se refleja en el control de los mecanismos de fosforilación. La piruvato quinasa y la fosfofructoquinasa II se inactivan por fosforilación dependiente de AMPc. Por lo tanto la fosforilación de la FF Quinasa II impide la formación de fructosa 2,6 di P, modulador alostérico positivo de la FF Quinasa I. Esto significa que cuando se incrementa el nivel de AMPc los niveles de fructosa 2,6 di P descienden y cesa la activación de la FF Quinasa I. La inducción enzimática de las enzimas FF quinasa I, piruvato quinasa y glucoquinasa por acción de la insulina es otro mecanismo de regulación. Es importante destacar que en muchos casos la inducción de proteínas - en este caso enzimas- también involucra la fosforilación de proteínas. Ciertas hormonas al unirse a su receptor desencadenan una cascada de fosforilaciones que impacta sobre factores de transcripción. Estos factores de transcripción, en su forma fosforilada, son capaces de promover la expresión de determinados genes. Por ejemplo la insulina, al unirse a su receptor, desencadena una serie de fosforilaciones que promueven la fosforilación de uno o más factores de transcripción involucrados en la expresión de genes que codifican por ejemplo para determinadas enzimas de la glucólisis. En algunos casos la fosforilación promovida por una hormona puede conducir a la fosforilación de un represor que regula la expresión de un determinado gen. La fosforilación de un represor determinado determina que éste pueda bloquear la transcripción de uno o más genes. Actualmente se conoce que el mecanismo de acción de hormonas que actúan vía la activación de receptores de membrana comprende la activación de una compleja red de moléculas de señalización, lo cual involucra la activación de diferentes quinasas. Un grupo de quinasas de reconocida participación en los mecanismos de señalización a través de membrana son las MAP quinasas. Por ejemplo se sabe que la unión del glucagon a su receptor de membrana no sólo promueve la activación de la proteína quinasa A (PKA), sino también la activación de la MAP quinasas ERK1/2. La insulina también activa miembros de la familia de estas enzimas. Las MAP quinasas (MAPK) (por ejemplo las quinasas ERK1/2) son serina/treonina quinasas que a su vez se activan por fosforilación en serina y tirosina, a través de una cascada de fosforilaciones en la que participan diferentes quinasas (ver esquema siguiente). Las MAP quinasas cumplen un papel muy importante en la transmisión de señales extracelulares al núcleo, promoviendo por ejemplo la expresión de genes. Asi por ejemplo un estímulo extracelular (Ej. Una hormona) puede activar una determinada MAP quinasa, la cual fosforila y activa uno o más factores de transcripción que participan en la expresión de un determinado gen. El concomitante aumento en los niveles del ARNm correspondiente lleva luego a un aumento en los niveles de la proteína correspondiente.La inducción de muchas de las enzimas de las vías de metabolización de la glucosa (por ejemplo glucólisis) por acción de insulina o glucagon se realiza a través de un mecanismo como el descripto, es decir con la participación de las MAPKs Balance energetico de la glucolisis. Formacion de ATP a partir del catabolismo de la glucosa: esto supone que el NADH formado en la glucólisis se transporta hacia las mitocondrias mediante el transbordador de malato. Si se usa el transbordador de glicerofosfato, sólo se formarán 1.5 ATP por cada mol de NADH. Note que hay una ventaja considerable en el uso de glucógeno en lugar de glucosa para la glucólisis anaerobia en el músculo, porque el producto de la glucógeno fosforilasa es la glucosa 1-fosfato, que es interconvertible con glucosa 6-fosfato. Esto ahorra el ATP que de otro modo sería usado por la hexocinasa, lo que aumenta el rendimiento neto de ATP de dos a tres por cada glucosa. Cuando un mol de glucosa es objeto de combustión en un calorímetro hasta CO2 y agua, se liberan unos 2 870 kJ como calor. Cuando la oxidación ocurre en los tejidos, se generan alrededor de 32 moles de ATP por cada molécula de glucosa oxidada hasta CO2 y agua. In vivo, la ΔG para la reacción de ATP sintasa se ha calculado como cercana a 51.6 kJ. Resulta que la energía total captada en el ATP por cada mol de glucosa oxidada es de 1 651 kJ, un 58% de la energía de combustión. La mayor parte del ATP se forma mediante fosforilación oxidativa originada por la reoxidación de coenzimas reducidas por la cadena respiratoria; el resto se forma por fosforilación a nivel de sustrato DATOS DE INTERÉS CLINICO RELACIONADOS CON LA GLUCÓLISIS CANCER Y GLUCÓLISIS Se conoce que en los tumores malignos están incrementadas las velocidades de captación de glucosa y de degradación de la glucosa por la vía glucolítica. Dado que las células cancerosas crecen más rapidamente que los vasos sanguíneos que las irriga, a medida que los tumores sólidos crecen, la disponibilidad de oxígeno se dificulta. Esto significa que el tejido experimenta hipoxia, por lo cual el metabolismo se torna fundamentalmente de tipo anaerobico. El estado de hipoxia favorece la activación de un factor de transcripción que se denomina HIF-1 (Hipoxia inducible transcription factor) el cual interviene en la inducción de transportadores de glucosa y de enzimas de la vía glucolítica. La inducción de estas enzimas es un mecanismo de adaptación que permite a la célula cancerosa sobrevivir en condiciones adversas (baja disponibilidad de oxígeno), hasta que se desarrolle la vascularización. Por otra parte, HIF 1 también participa en la expresión del factor de crecimiento endotelial de vasos (VEGF, vascular endotelial growth factor), que es fundamental para el desarrollo de los vasos sanguíneos. El desarrollo de fármacos apunta a producir fármacos que impidan la vascularización para evitar asi el desarrollo del tumor. ACIDOSIS LÁCTICA La acidosis láctica se presenta cuando los niveles de lactato en sangre son superiores a 5mM y el pH sanguíneo está por debajo del valor normal. Puede darse por una sobreproducción de lactato o por una disminución en el consumo de lactato o por ambas causas. La causa más común de la acidosis láctica es una producción exacerbada de lactato, por ejemplo durante el ejercicio. Como ya discutimos, el lactato se produce por glucólisis anaeróbica. Cuando la oxidación de la glucosa ocurre por glucólisis anaeróbica, el rendimiento energético es mucho menor que el obtenido por glucólisis aeróbica. Esto significa que para generar la energía necesaria, el organismo debe incrementar la velocidad de consumo de glucosa, y por ende la producción de lactato. Además, el hecho que en condiciones anaeróbicas está disminuído el consumo de lactato incrementa aún más la concentración de lactato en sangre. Esto se explica porque los dos procesos que consumen el lactato, es decir su oxidación a CO2 y H2O o bien su reconversión a glucosa, ambos procesos requieren oxígeno. La oxigenación de los tejidos permite controlar este tipo de acidosis. ANEMIA HEMOLÍTICA POR DÉFICIT DE PIRUVATO QUINASA La producción de ATP en los eritrocitos maduros depende exclusivamente de la vía glucolítica. Cuando la producción de ATP no es suficiente, ciertas actividades enzimàticas se alteran, en particular las bombas iónicas como Na/K ATPasa. La actividad de esta bomba es importante porque contribuye a mantener la forma del eritrocito, permitiendo su deslizamiento por los capilares sin llegar a romperse. Esto explica porque cuando no hay suficiente producción de ATP en el eritrocito éste se torna frágil y se rompe fácilmente. La anemia que caracteriza a este cuadro se denomina anemia hemolítica. Ciertos pacientes tienen una actividad deficiente de la enzima piruvato quinasa. Los afectados tienen una actividad de piruvato quinasa en eritrocitos que suele ser sólo del 25% del valor registrado en individuos sanos. Esta falla enzimática es poco frecuente, pero su ocurrencia está asociada con anemia hemolítica. Bueno gente eso es todo!!! Espero les sea de mucha utilidad y que ante cualquier duda pueden consultar. Hasta el proximo post...Saludos!!!

Hola Taringeros, como les va??? Hoy les quiero comentar algunas cositas sobre esta enfermedad infecciosa que tanto se hace nombrar en estos dias!!! ¿QUE ES LA GRIPE? La gripe es una enfermedad producida por un virus (llamado influenza en el mundo científico). Este virus es capaz de cambiar año a año, lo cual hace que al organismo le resulte muy difícil establecer una defensa eficaz contra el mismo. Cuando se padece gripe, la sintomatología es muy evidente: en los seres humanos (tambien afecta a avesy otros mamiferos) afecta a las vías respiratorias, inicialmente puede ser similar a un resfriado y con frecuencia se acompaña de síntomas generales como dolor de garganta, debilidad, dolores musculares (mialgias), dolor estomacal, articulares (artralgias) y de cabeza (cefalea), con tos (que generalmente es seca y sin mucosidad), malestar general y algunos signos como pueda ser la fiebre. En algunos casos más graves puede complicarse con pulmonía (neumonía), que puede resultar mortal, especialmente en niños pequeños y sobre todo en ancianos. Aunque se puede confundir con el resfriado común. La gripe se transmite desde individuos infectados a través de gotas en aerosol cargadas de virus (procedentes de saliva, secreción nasal y bronquial), que son emitidas con la tos o los estornudos o sólo al hablar. Ya mucho más raramente, a través de las heces de pájaros infectados. También es transmisible por la sangre y por las superficies u objetos contaminados con el virus, que se denominan fomites. UN POCO DE HISTORIA... Los síntomas de la gripe en humanos fueron descritos por Hipócrates hace unos 2.400 años.Desde entonces el virus ha causado, además de la epidemia anual, numerosas pandemias(epidemia global). La pandemia más letal y conocida fue la denominada gripe española (virus A, subtipo H1N1), que duró desde 1918 a 1919. Se denomina así porque España era el país que publicaba más datos sobre el desastre sanitario que estaba ocasionando pues muchos los censuraban con motivo de la primera guerra mundial en la cual estaban inmersos. Las estimaciones más antiguas hablaban de unos 40 ó 50 millones de muertos causados por ella,aunque las aproximaciones más actuales sitúan la cifra entre 50 y 100 millones de personas fallecidas por aquella gripe en todo el mundo.Esta pandemia ha sido descrita como el mayor holocausto médico de la historia, y causó al menos tantos muertos como la peste negra.Esta gran mortalidad fue debida a la gran tasa de infectividad (hasta el 50% de la población expuesta) y a la gravedad de los síntomas causados por la producción masiva de citoquinas (tormenta de citoquinas). La familia de virus Orthomyxoviridae es la causante (etiología) de la gripe, y fue descrita por primera vez en cerdos por Richard Schope en 1931.Este descubrimiento fue seguido en breve por el aislamiento del virus en humanos por un grupo de investigación dirigido por Patrick Laidlaw y el Medical Research Council del Reino Unido en 1933Sin embargo, hubo que esperar hasta 1935 para que Wendell Meredith Stanley, Premio Nobel de Química 1946, estableciera la verdadera naturaleza no celular de los virus. ETIOLOGIA DE LA GRIPE...Es decir ¿Cual es su Causa? Los virus causantes de la gripe en el hombre, se clasifican en tres tipos influenza similares morfológicamente: A, B y C. Los virus de la gripe A, la más frecuente y estudiada, son los más importantes por su capacidad de sufrir cambios antigénicos, dando lugar epidemias y pandemias e infectar a poblaciones animales.El virus A, se subtipifica según dos antígenos de superficie: hemaglutinina (H) y neuraminidasa (N), dando lugar esa denominación que tan familiar resulta hoy en los medios de comunicación con el asunto de la gripe aviar. La identificación completa de la cepa contiene: el lugar de origen, número de aislamiento, año de aislamiento y subtipo (p. ej. virus gripal A/Victoria/3/79 H3N2).Los virus de la gripe B y C se identifican de forma similar, pero los antígenos H y N no tienen denominaciones de subtipo al haber menos variacion. La inmunidad contra el virus gripal la representan los anticuerpos contra el antígeno H, en tanto que los anticuerpos contra el antígeno N reducen la propagación del virus. AQUI ESTA EL CULPABLE... El virus de la gripe tiene una estructura relativamente simple. Una envuelta lipídica envuelve el caparazón proteico o cápsida, el cual encierra el material genético enrollado. Desde esta envuelta se proyectan dos tipos de proteínas a modo de púas, la hemoaglutinina y la neuraminidasa. Estas dos proteínas actúan como antígenos, provocando una respuesta inmune en el organismo invadido por el virus. El virus de la gripe tiene la capacidad de mutar periódicamente estas proteínas. De esta manera, los hospedadores humanos deben producir nuevas defensas inmunes cada vez que las proteínas mutan; de aquí las vacunaciones anuales que se realizan. PATOGENIA Y PATOLOGIA... Los virus de la gripe se fijan mediante hemaglutininas a los azúcares de ácido siálico de la membrana celular de las células epiteliales mucosas de las fosas nasales, garganta y pulmones (más en nasofaringe y tráquea pues la afectación bronquiolar y alveolar se da sólo en los casos graves), en los mamíferos, y del intestino, en las aves. Una explicacion muy sencilla de como actua... TRATAMIENTO... Hay 2 tipos: Sintomático: Los consejos generales para una persona afectada de gripe son reposo, ingesta abundante de líquidos, evitar el consumo de alcohol (licor) y tabaco (en el caso improbable de que le apetezca al paciente, al menos los primeros días) y un fármaco que alivie los síntomas, como el paracetamol, que siendo antitérmico y también analgésico alivia simultáneamente la fiebre y el dolor. Se puede asociar con codeína (salvo en pacientes respiratorios crónicos pues deprime el centro respiratorio) para así evitar la tos pues es improductiva y por lo tanto su supresión no es problemática ya que no provoca acumulo de secreciones. La asociación comercial de paracetamol con codeína no sirve pues la dosis de ésta sólo tiene efecto analgésico y no antitusígeno. La aspirina no está indicada en niños y adolescentes (y el tratamiento previo parece aumentar la susceptibilidad de padecerla) con síntomas de gripe (y tampoco en cualquier otra situación febril) para evitar la aparición del síndrome de Reye, una complicación infrecuente pero grave del hígado y del cerebro que puede afectarles cuando toman este antiinflamatorio en el contexto de algunas enfermedades víricas (especialmente la infección por Influenzavirus B). Preventivo Antiviral: En el caso de la gripe existe una vacuna preventiva recomendable. Todos los años, la Organización Mundial de la Salud recomienda la composición de la vacuna a utilizar en aquellas personas incluidas en los grupos de riesgo, a personas ancianas, ciertos enfermos, grupos que puedan transmitir la enfermedad y aquellos que trabajen en ciertos servicios públicos. Bueno taringueros eso es todo!!!Seguramente habra correcciones que hacer pero bueno es solo para darles un poco de informacion sobre, como ya se habran dado cuenta jajaja, la gripe... Saludos y muchas gracias por visitar el post...Espero les sea de utilidad.